

Unravelling cell migration: defining movement from the cell surface
- PMID: 35499121
- PMCID: PMC9067518
- DOI: 10.1080/19336918.2022.2055520
Unravelling cell migration: defining movement from the cell surface
Abstract
Cell motility is essential for life and development. Unfortunately, cell migration is also linked to several pathological processes, such as cancer metastasis. Cells' ability to migrate relies on many actors. Cells change their migratory strategy based on their phenotype and the properties of the surrounding microenvironment. Cell migration is, therefore, an extremely complex phenomenon. Researchers have investigated cell motility for more than a century. Recent discoveries have uncovered some of the mysteries associated with the mechanisms involved in cell migration, such as intracellular signaling and cell mechanics. These findings involve different players, including transmembrane receptors, adhesive complexes, cytoskeletal components , the nucleus, and the extracellular matrix. This review aims to give a global overview of our current understanding of cell migration.
Keywords: Cell migration; cell mechanics; extracellular matrix.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
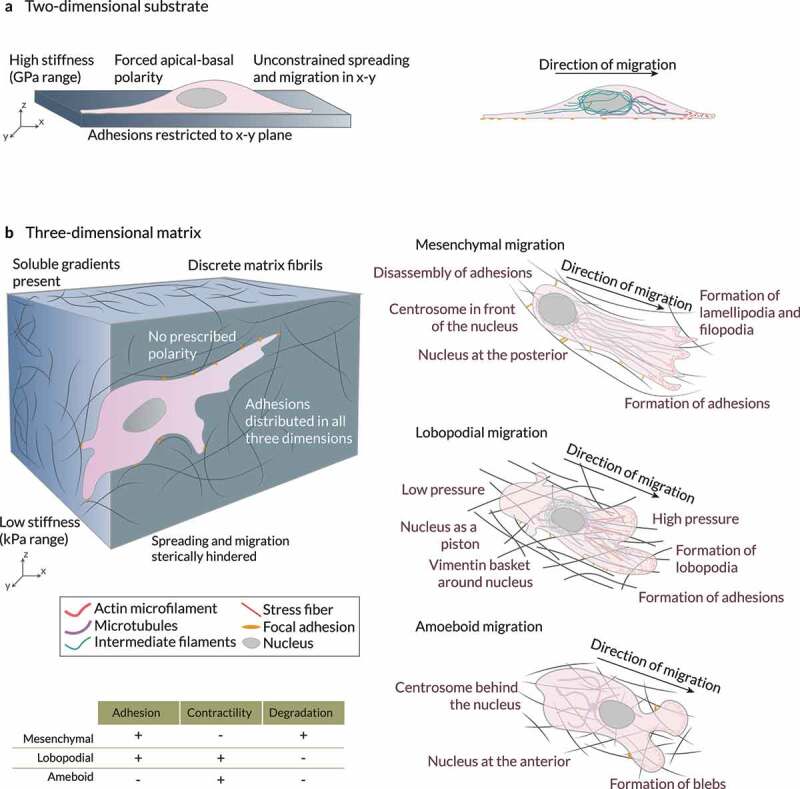
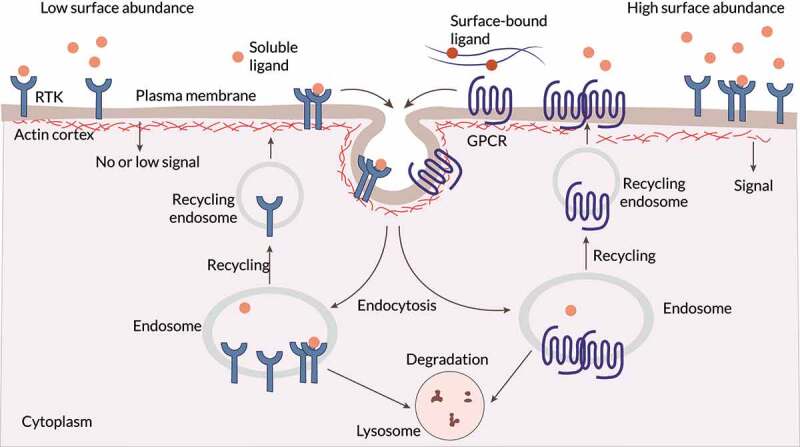
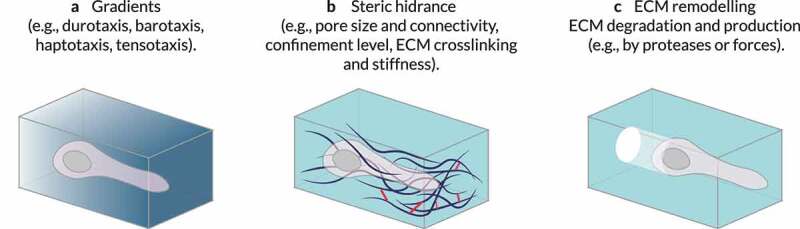
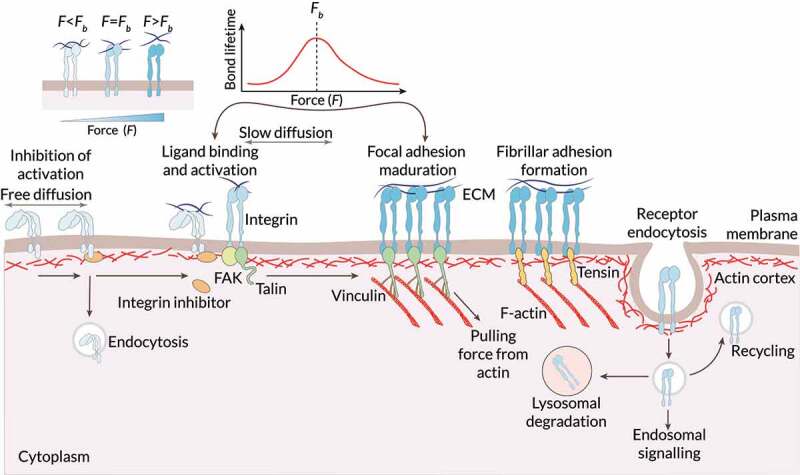
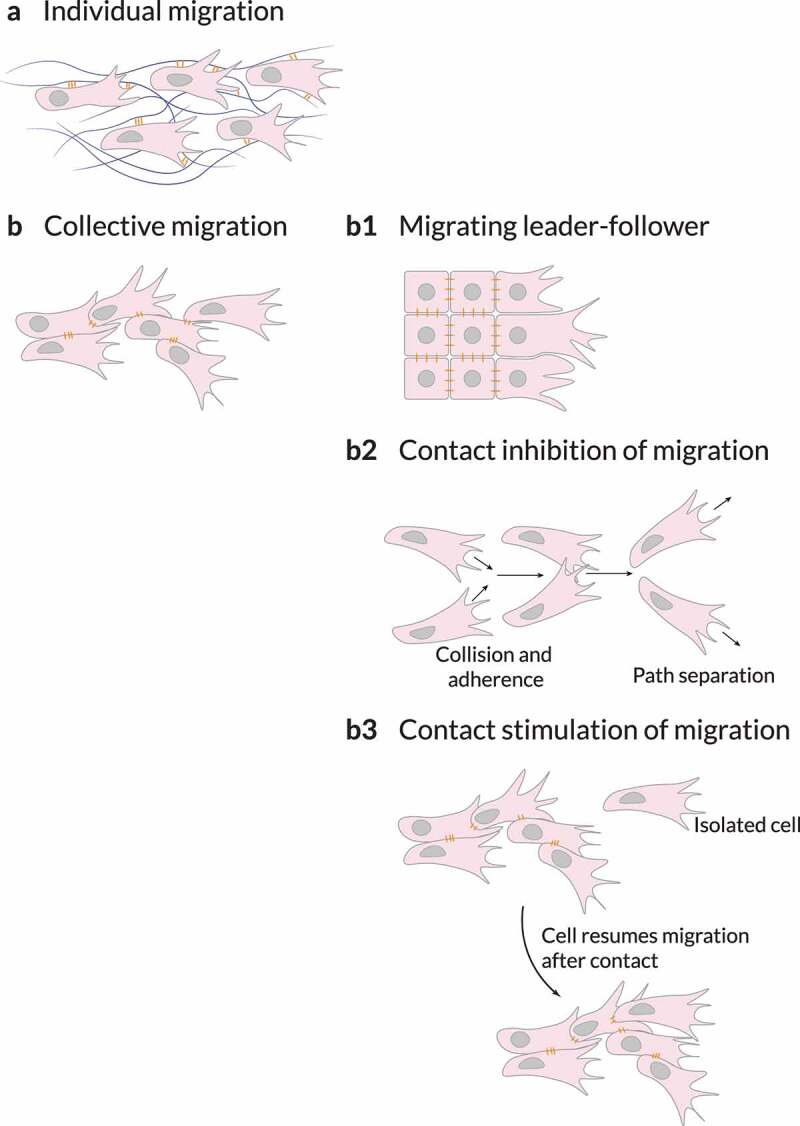
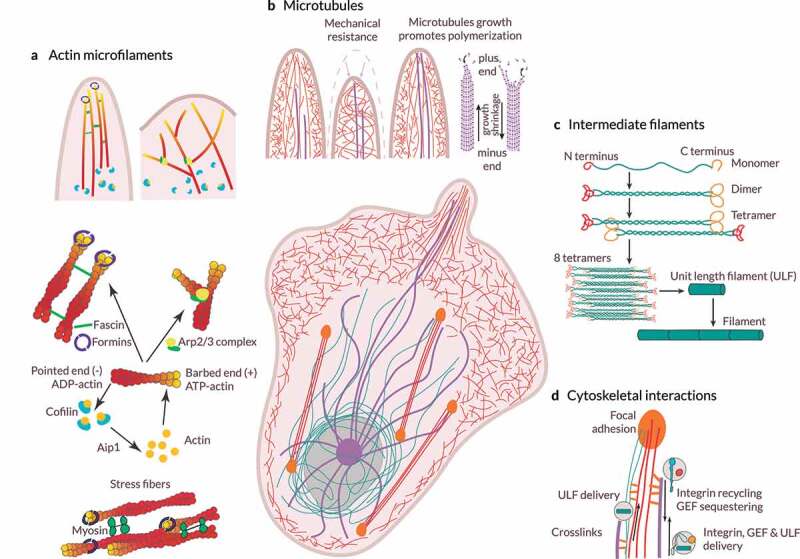
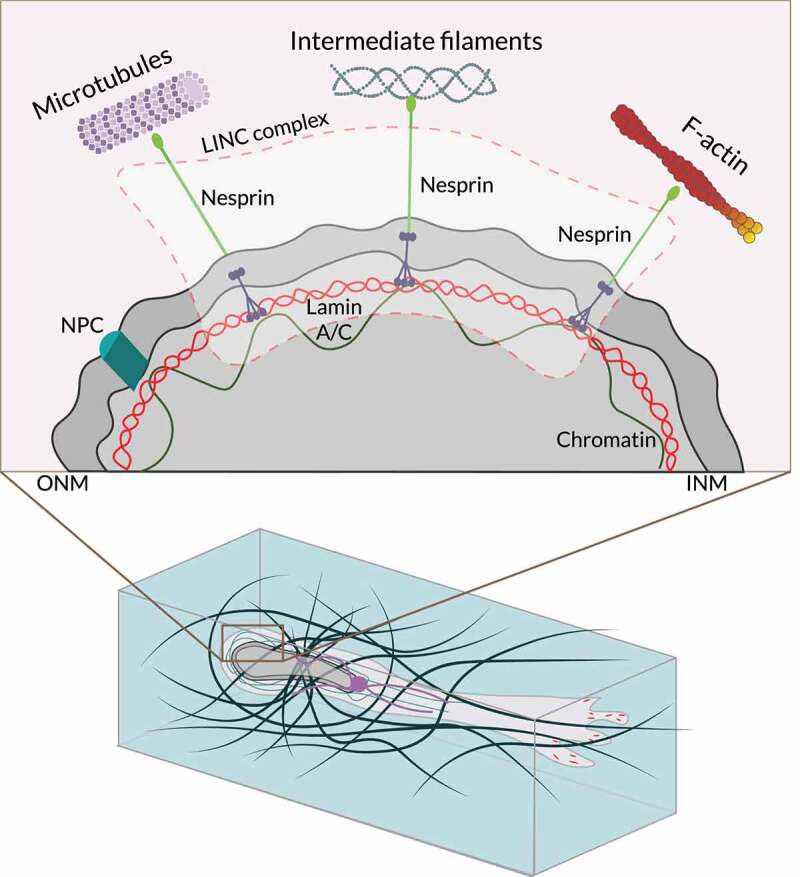
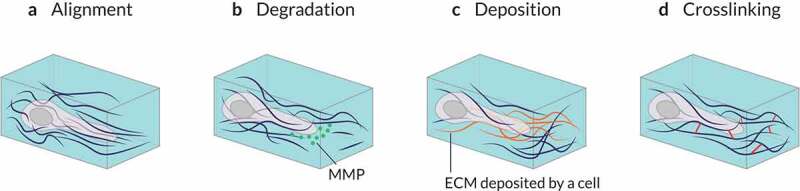
References
-
- Karamanos NK, Theocharis AD, Piperigkou Z, et al. A guide to the composition and functions of the extracellular matrix. FEBS J. 2021;228:15776. - PubMed
-
- Marsha CL, Cynthia AR-K.. Targeting extracellular matrix stiffness to attenuate disease: from molecular mechanisms to clinical trials. Sci Transl Med. 10(422):2018. - PubMed
-
- Kenneth MY, Sixt M. Mechanisms of 3D cell migration. Nat Rev Mol Cell Biol. 2019;20(12):738–752. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources