

The endoplasmic reticulum membrane protein complex localizes to the mitochondrial - endoplasmic reticulum interface and its subunits modulate phospholipid biosynthesis in Trypanosoma brucei
- PMID: 35500022
- PMCID: PMC9113592
- DOI: 10.1371/journal.ppat.1009717
The endoplasmic reticulum membrane protein complex localizes to the mitochondrial - endoplasmic reticulum interface and its subunits modulate phospholipid biosynthesis in Trypanosoma brucei
Abstract
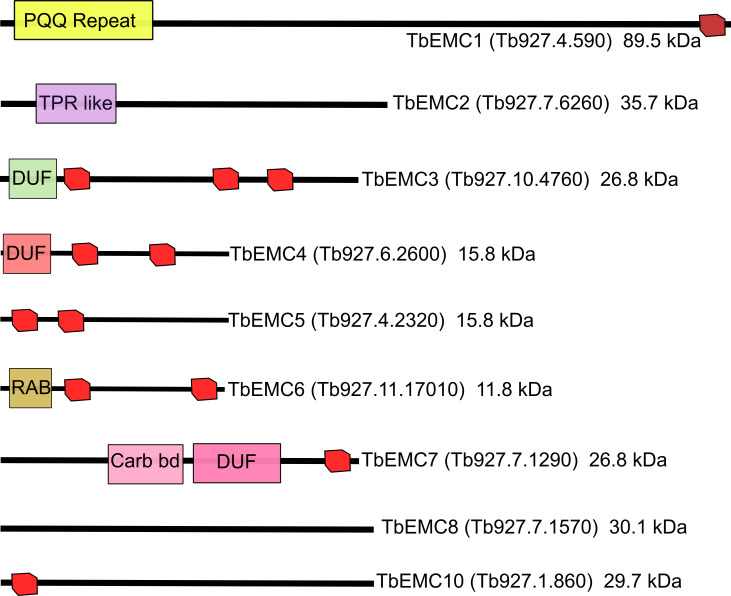
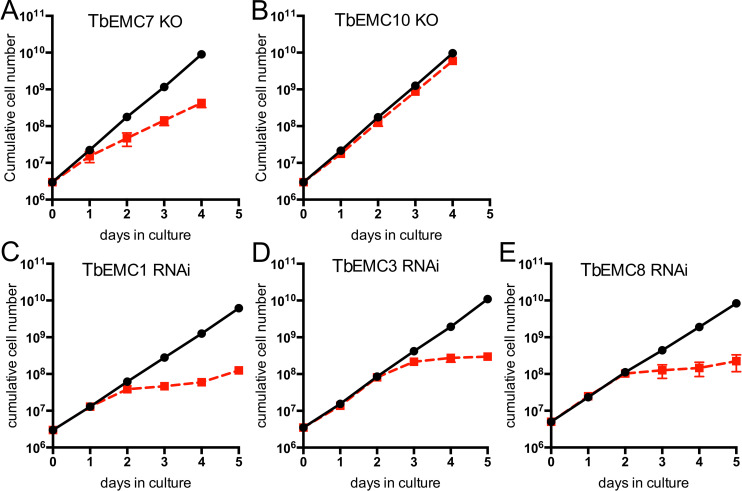
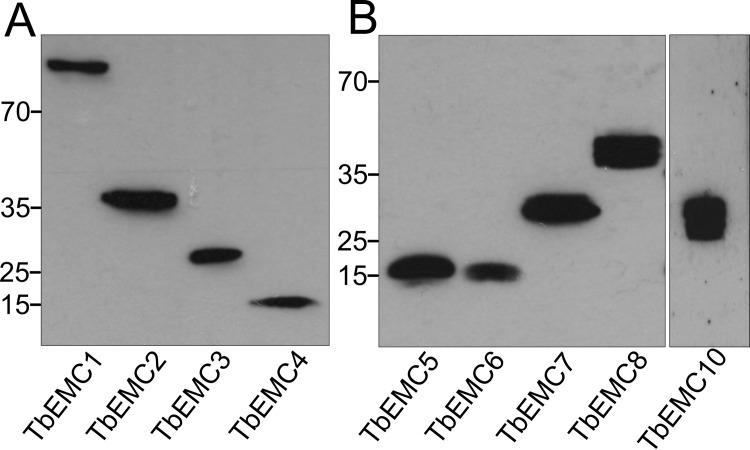
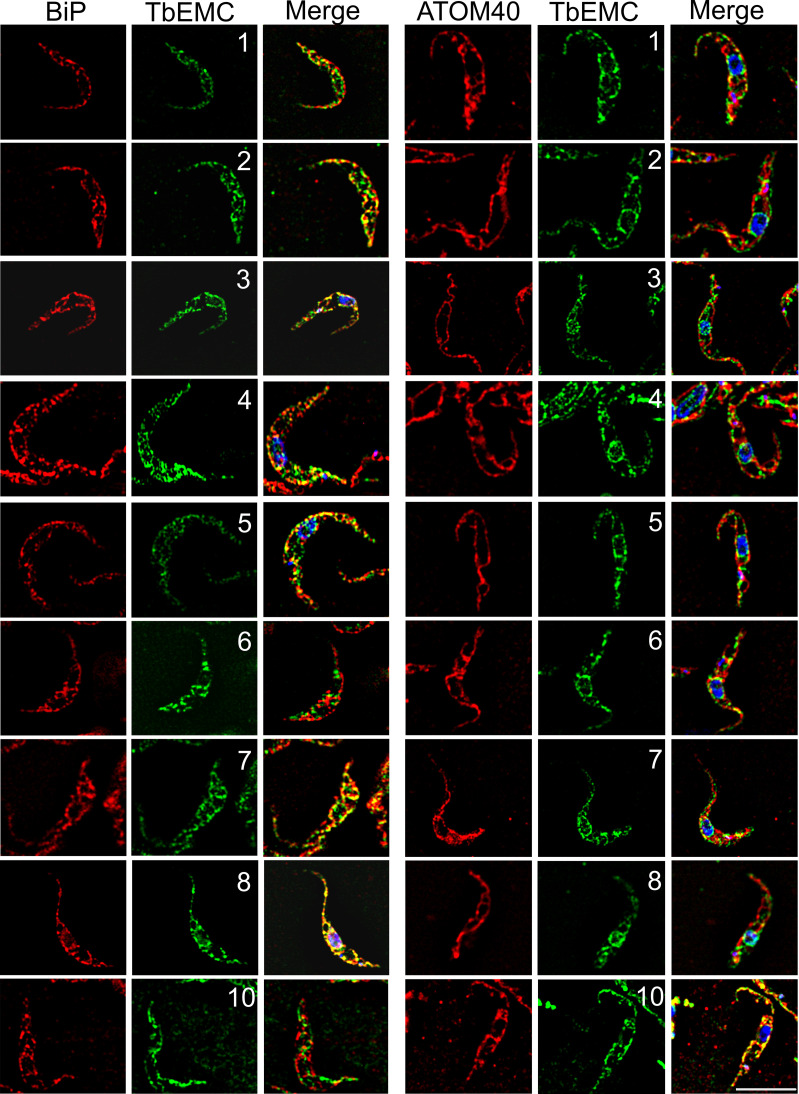
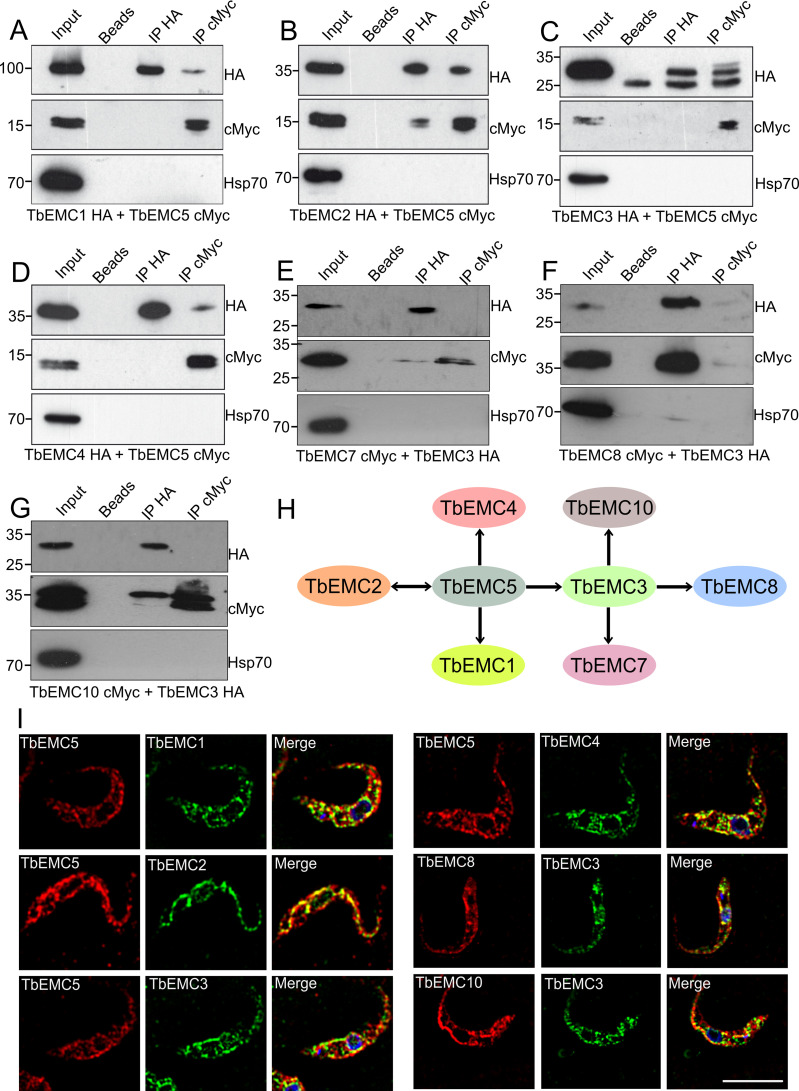
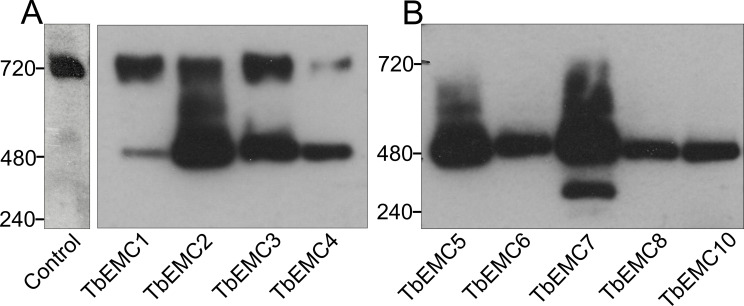
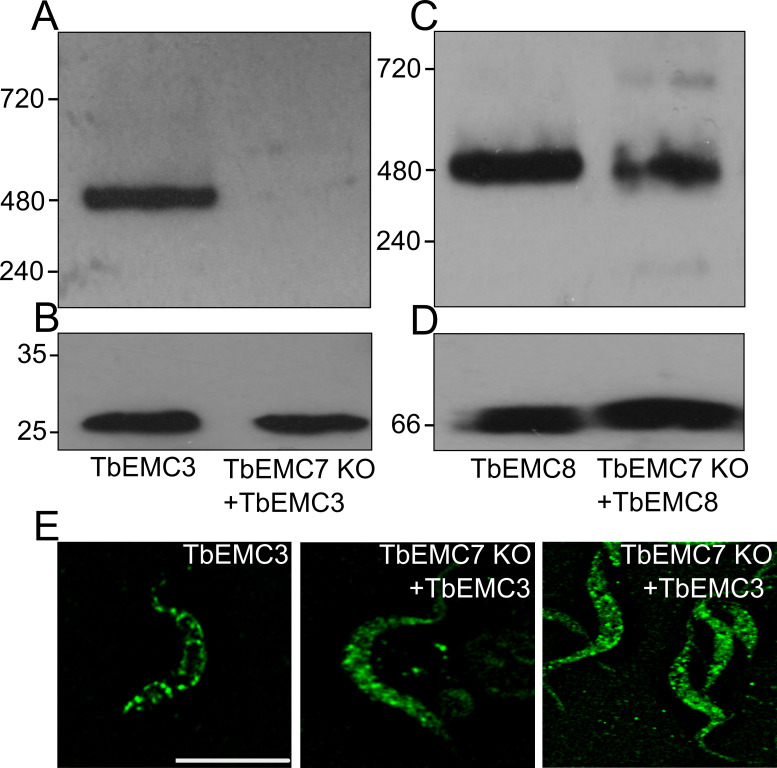
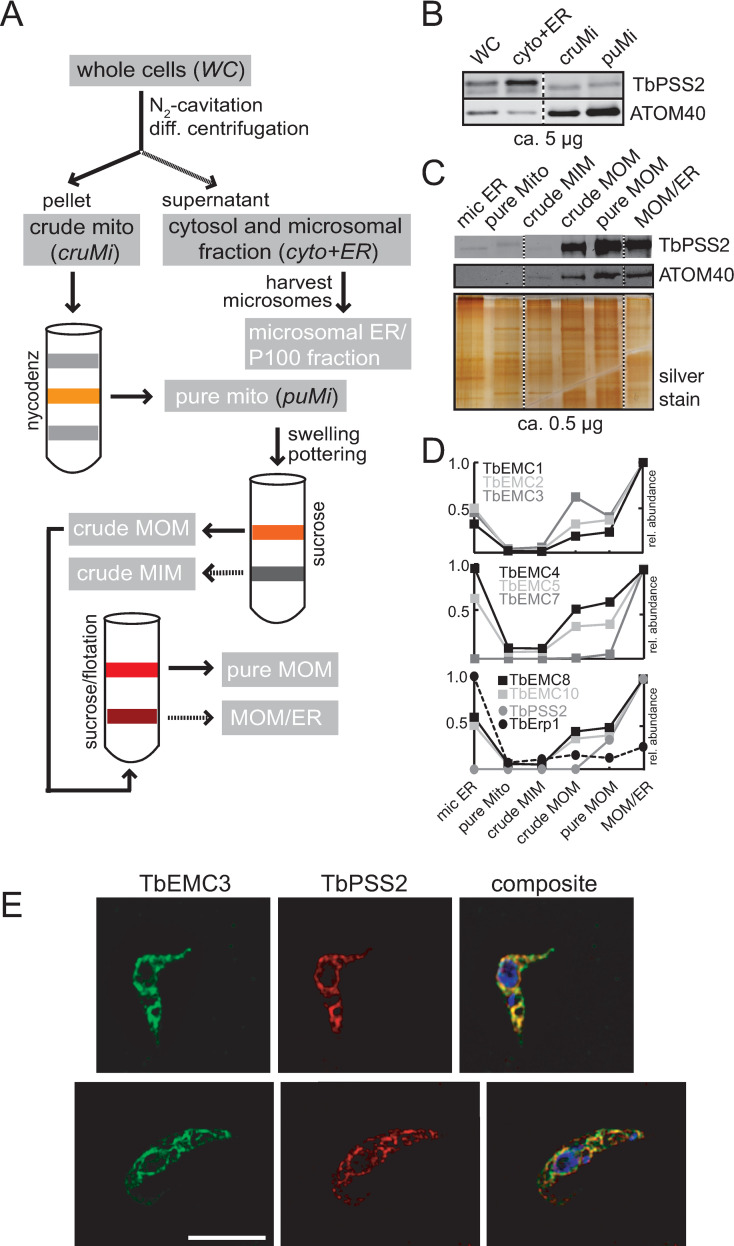
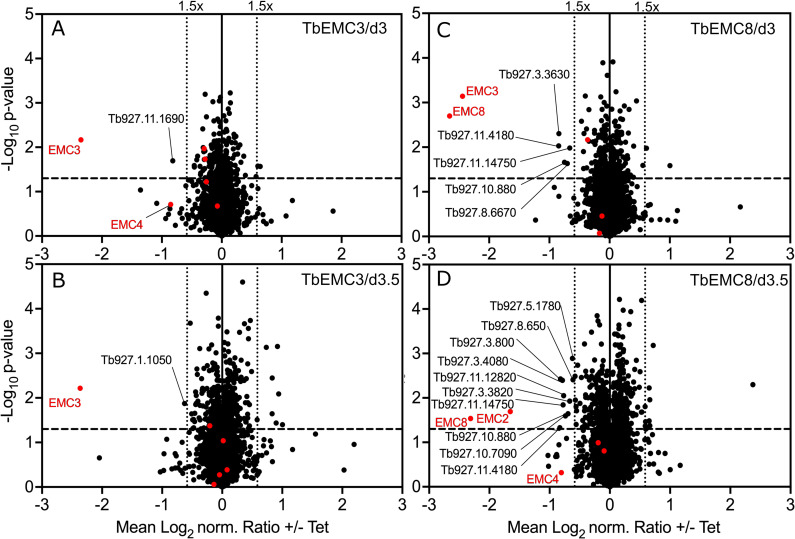
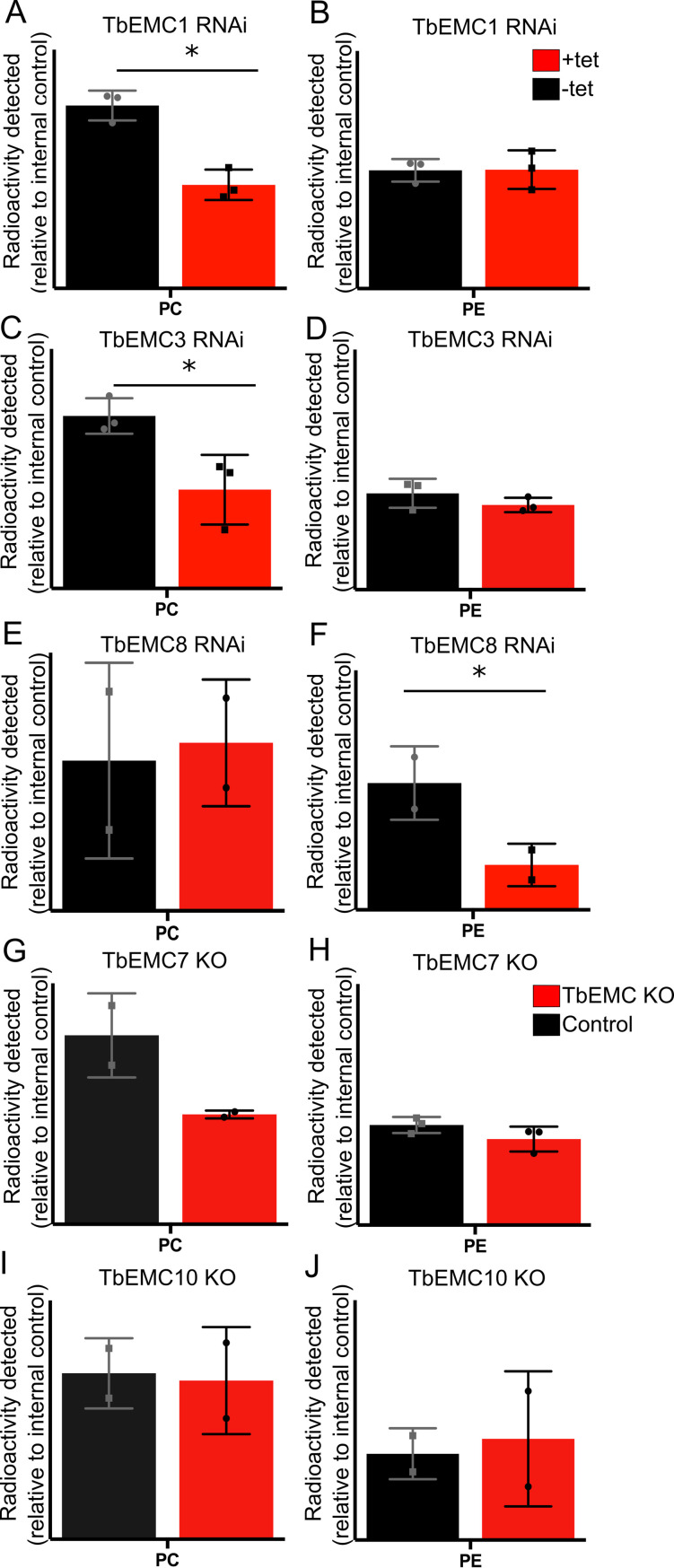
The endoplasmic reticulum membrane complex (EMC) is a versatile complex that plays a key role in membrane protein biogenesis in the ER. Deletion of the complex has wide-ranging consequences including ER stress, disturbance in lipid transport and organelle tethering, among others. Here we report the function and organization of the evolutionarily conserved EMC (TbEMC) in the highly diverged eukaryote, Trypanosoma brucei. Using (co-) immunoprecipitation experiments in combination with mass spectrometry and whole cell proteomic analyses of parasites after depletion of select TbEMC subunits, we demonstrate that the TbEMC is composed of 9 subunits that are present in a high molecular mass complex localizing to the mitochondrial-endoplasmic reticulum interface. Knocking out or knocking down of single TbEMC subunits led to growth defects of T. brucei procyclic forms in culture. Interestingly, we found that depletion of individual TbEMC subunits lead to disruption of de novo synthesis of phosphatidylcholine (PC) or phosphatidylethanolamine (PE), the two most abundant phospholipid classes in T. brucei. Downregulation of TbEMC1 or TbEMC3 inhibited formation of PC while depletion of TbEMC8 inhibited PE synthesis, pointing to a role of the TbEMC in phospholipid synthesis. In addition, we found that in TbEMC7 knock-out parasites, TbEMC3 is released from the complex, implying that TbEMC7 is essential for the formation or the maintenance of the TbEMC.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Identification and functional characterization of Trypanosoma brucei peroxin 16.Biochim Biophys Acta. 2015 Oct;1853(10 Pt A):2326-37. doi: 10.1016/j.bbamcr.2015.05.024. Epub 2015 May 27. Biochim Biophys Acta. 2015. PMID: 26025675
-
Phosphatidylethanolamine and phosphatidylcholine biosynthesis by the Kennedy pathway occurs at different sites in Trypanosoma brucei.Sci Rep. 2015 Nov 18;5:16787. doi: 10.1038/srep16787. Sci Rep. 2015. PMID: 26577437 Free PMC article.
-
Localization of a Trypanosome Peroxin to the Endoplasmic Reticulum.J Eukaryot Microbiol. 2017 Jan;64(1):97-105. doi: 10.1111/jeu.12343. Epub 2016 Jul 18. J Eukaryot Microbiol. 2017. PMID: 27339640 Free PMC article.
-
The role of phospholipids in the biological activity and structure of the endoplasmic reticulum.Biochim Biophys Acta. 2013 Nov;1833(11):2499-510. doi: 10.1016/j.bbamcr.2013.05.018. Epub 2013 May 24. Biochim Biophys Acta. 2013. PMID: 23711956 Review.
-
The Role of EMC during Membrane Protein Biogenesis.Trends Cell Biol. 2019 May;29(5):371-384. doi: 10.1016/j.tcb.2019.01.007. Epub 2019 Feb 27. Trends Cell Biol. 2019. PMID: 30826214 Review.
Cited by
-
Roles for PMP22 in Schwann cell cholesterol homeostasis in health and disease.Biochem Soc Trans. 2024 Aug 28;52(4):1747-1756. doi: 10.1042/BST20231359. Biochem Soc Trans. 2024. PMID: 38979632 Free PMC article. Review.
-
How host ER membrane chaperones and morphogenic proteins support virus infection.J Cell Sci. 2023 Jul 1;136(13):jcs261121. doi: 10.1242/jcs.261121. Epub 2023 Jul 4. J Cell Sci. 2023. PMID: 37401530 Free PMC article.
-
The endoplasmic reticulum of trypanosomatids: An unrevealed road for chemotherapy.Front Cell Infect Microbiol. 2022 Nov 10;12:1057774. doi: 10.3389/fcimb.2022.1057774. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36439218 Free PMC article. Review.
-
Protein control of membrane and organelle dynamics: Insights from the divergent eukaryote Toxoplasma gondii.Curr Opin Cell Biol. 2022 Jun;76:102085. doi: 10.1016/j.ceb.2022.102085. Epub 2022 May 12. Curr Opin Cell Biol. 2022. PMID: 35569259 Free PMC article. Review.
