

Sound Localization of World and Head-Centered Space in Ferrets
- PMID: 35501154
- PMCID: PMC7612817
- DOI: 10.1523/JNEUROSCI.0291-22.2022
Sound Localization of World and Head-Centered Space in Ferrets
Abstract
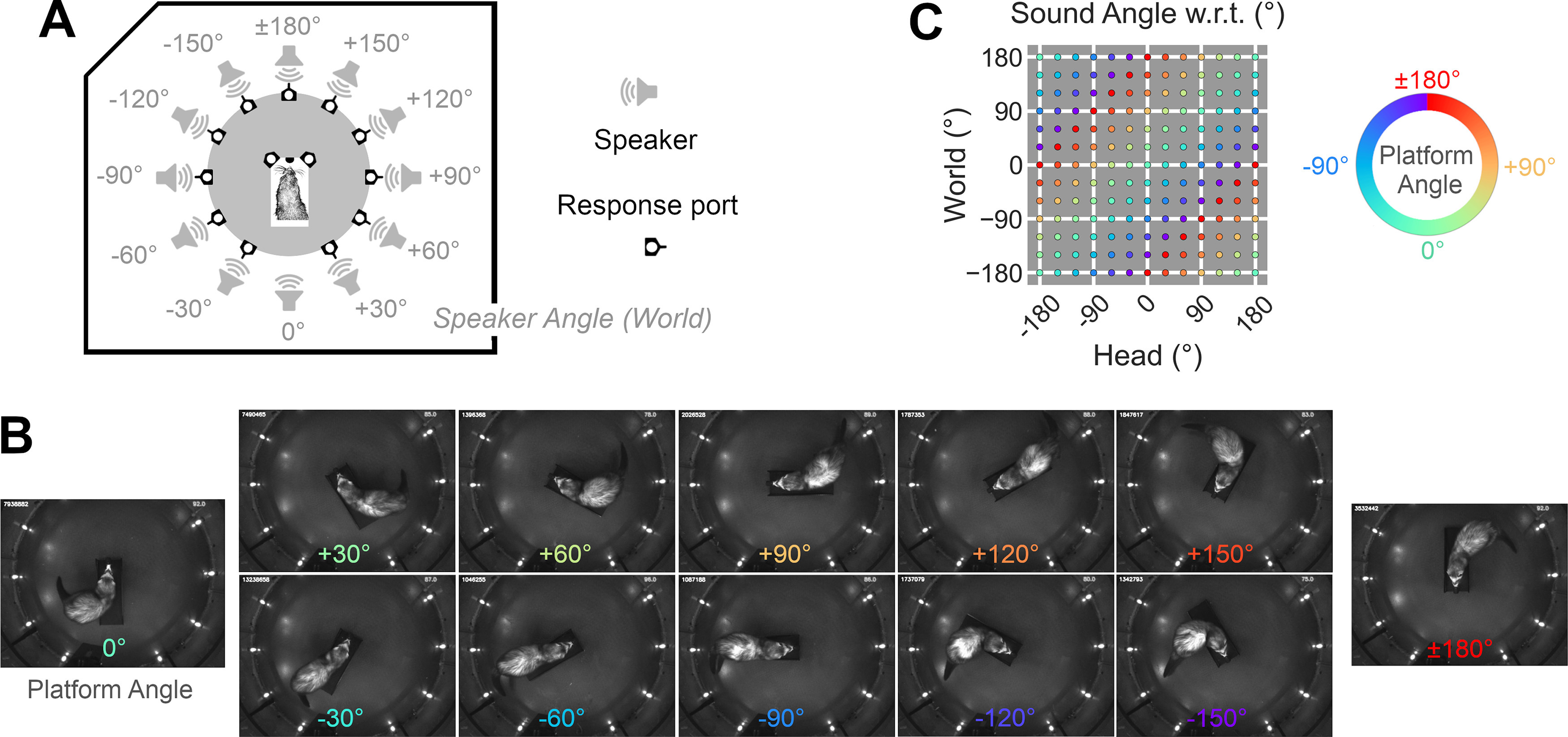
The location of sounds can be described in multiple coordinate systems that are defined relative to ourselves, or the world around us. Evidence from neural recordings in animals point toward the existence of both head-centered and world-centered representations of sound location in the brain; however, it is unclear whether such neural representations have perceptual correlates in the sound localization abilities of nonhuman listeners. Here, we establish novel behavioral tests to determine the coordinate systems in which ferrets can localize sounds. We found that ferrets could learn to discriminate between sound locations that were fixed in either world-centered or head-centered space, across wide variations in sound location in the alternative coordinate system. Using probe sounds to assess broader generalization of spatial hearing, we demonstrated that in both head and world-centered tasks, animals used continuous maps of auditory space to guide behavior. Single trial responses of individual animals were sufficiently informative that we could then model sound localization using speaker position in specific coordinate systems and accurately predict ferrets' actions in held-out data. Our results demonstrate that ferrets, an animal model in which neurons are known to be tuned to sound location in egocentric and allocentric reference frames, can also localize sounds in multiple head and world-centered spaces.SIGNIFICANCE STATEMENT Humans can describe the location of sounds either relative to themselves, or in the world, independent of their momentary position. These different spaces are also represented in the activity of neurons in animals, but it is not clear whether nonhuman listeners also perceive both head and world-centered sound location. Here, we designed behavioral tasks in which ferrets discriminated between sounds using their position in the world, or relative to the head. Subjects learnt to solve both problems and generalized sound location in each space when presented with infrequent probe sounds. These findings reveal a perceptual correlate of neural sensitivity previously observed in the ferret brain and establish that, like humans, ferrets can access an auditory map of their local environment.
Keywords: auditory; behavior; ferret; hearing; modeling; sound localization.
Copyright © 2022 the authors.
Figures







References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources