

Neuroplasticity related to chronic pain and its modulation by microglia
- PMID: 35501933
- PMCID: PMC9063368
- DOI: 10.1186/s41232-022-00199-6
Neuroplasticity related to chronic pain and its modulation by microglia
Abstract
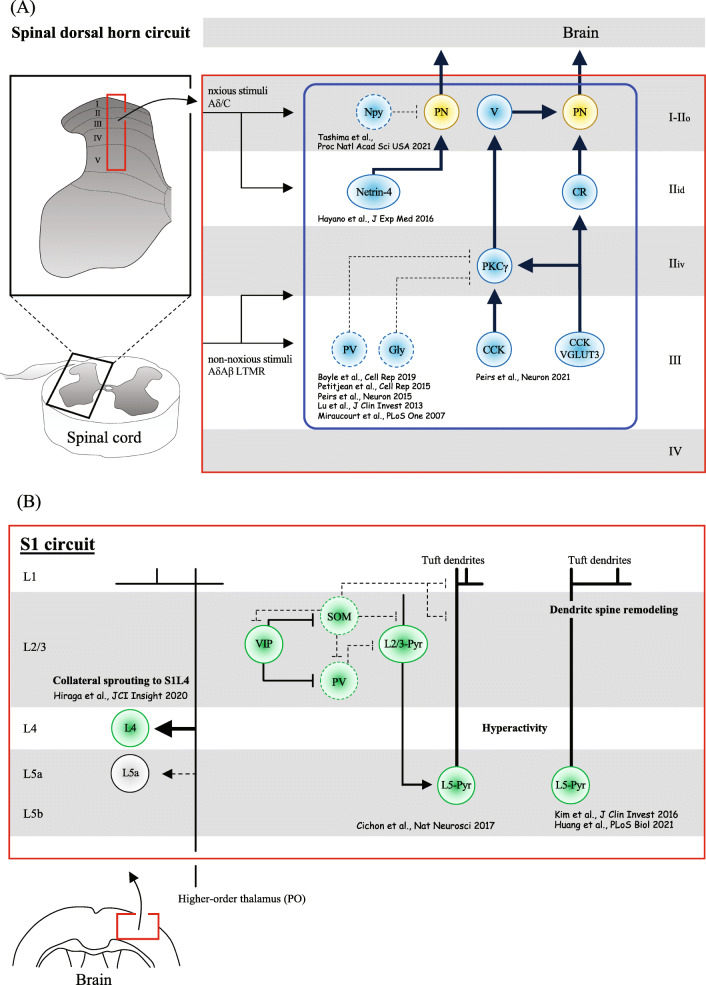
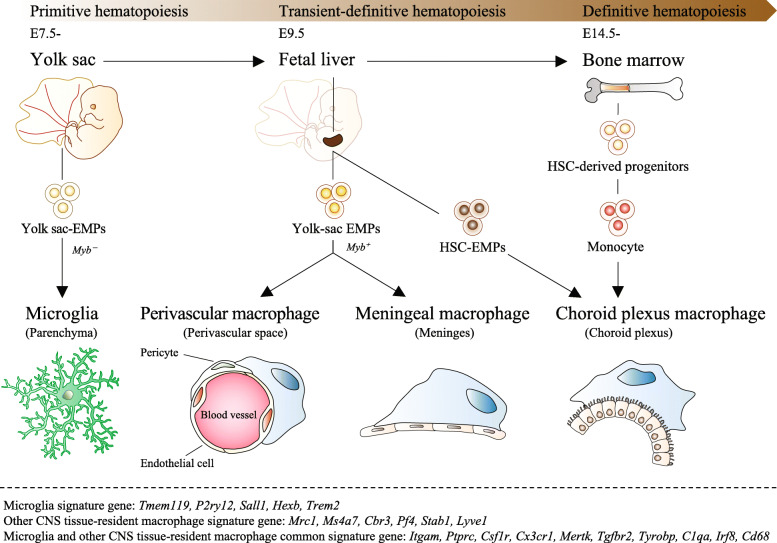
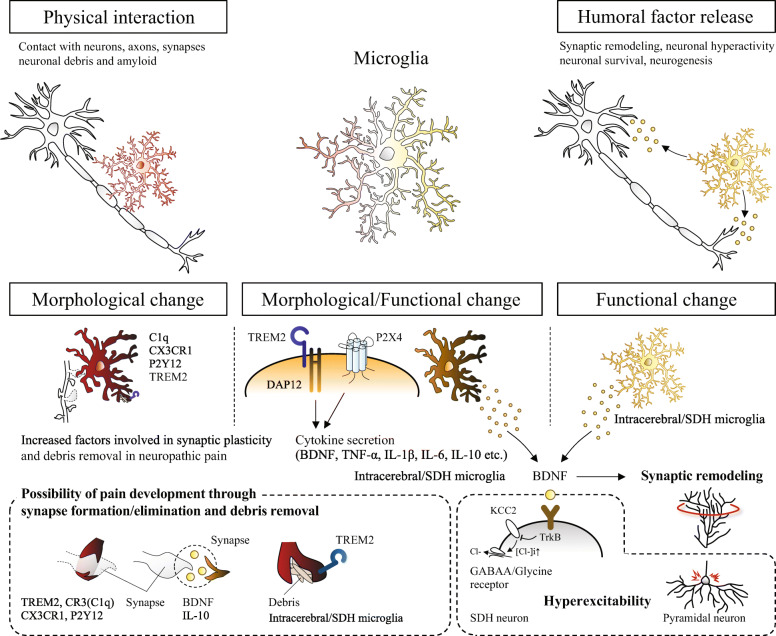
Neuropathic pain is often chronic and can persist after overt tissue damage heals, suggesting that its underlying mechanism involves the alteration of neuronal function. Such an alteration can be a direct consequence of nerve damage or a result of neuroplasticity secondary to the damage to tissues or to neurons. Recent studies have shown that neuroplasticity is linked to causing neuropathic pain in response to nerve damage, which may occur adjacent to or remotely from the site of injury. Furthermore, studies have revealed that neuroplasticity relevant to chronic pain is modulated by microglia, resident immune cells of the central nervous system (CNS). Microglia may directly contribute to synaptic remodeling and altering pain circuits, or indirectly contribute to neuroplasticity through property changes, including the secretion of growth factors. We herein highlight the mechanisms underlying neuroplasticity that occur in the somatosensory circuit of the spinal dorsal horn, thalamus, and cortex associated with chronic pain following injury to the peripheral nervous system (PNS) or CNS. We also discuss the dynamic functions of microglia in shaping neuroplasticity related to chronic pain. We suggest further understanding of post-injury ectopic plasticity in the somatosensory circuits may shed light on the differential mechanisms underlying nociceptive, neuropathic, and nociplastic-type pain. While one of the prominent roles played by microglia appears to be the modulation of post-injury neuroplasticity. Therefore, future molecular- or genetics-based studies that address microglia-mediated post-injury neuroplasticity may contribute to the development of novel therapies for chronic pain.
Keywords: CNS injury; Microglia; Neuroplasticity; Nociplastic pain; PNS injury; Thalamic hemorrhage.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
