

Simultaneous stabilization of actin cytoskeleton in multiple nephron-specific cells protects the kidney from diverse injury
- PMID: 35504916
- PMCID: PMC9065033
- DOI: 10.1038/s41467-022-30101-4
Simultaneous stabilization of actin cytoskeleton in multiple nephron-specific cells protects the kidney from diverse injury
Abstract
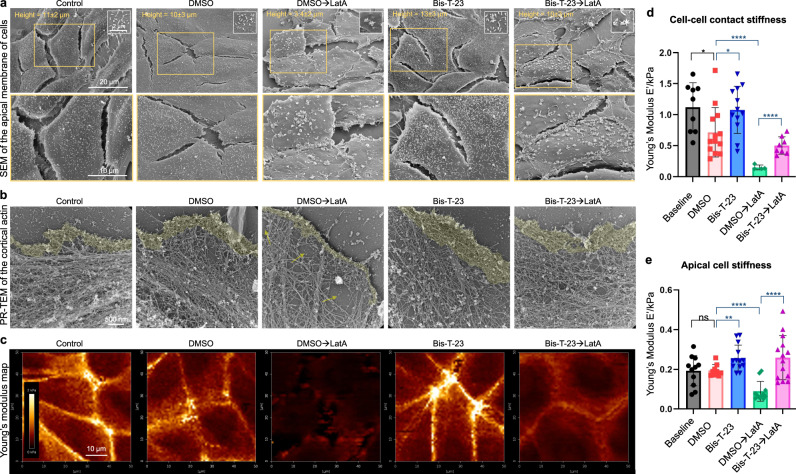
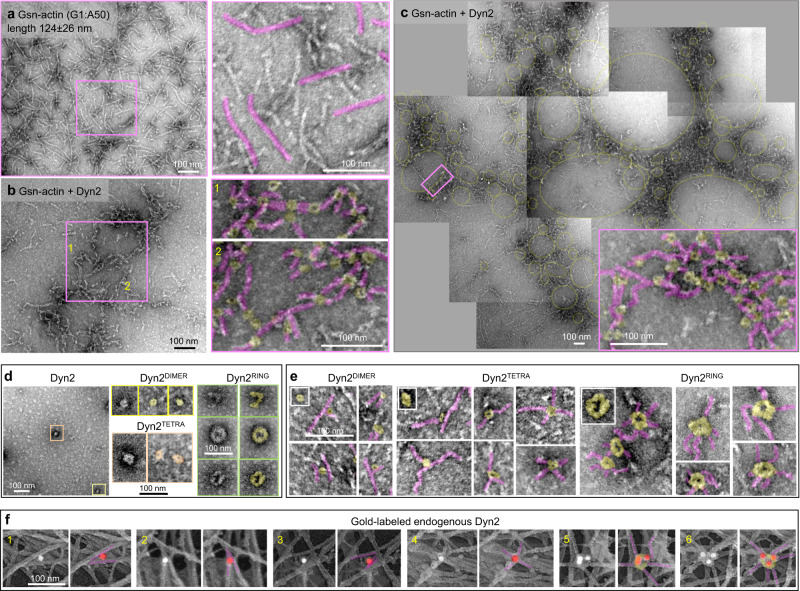
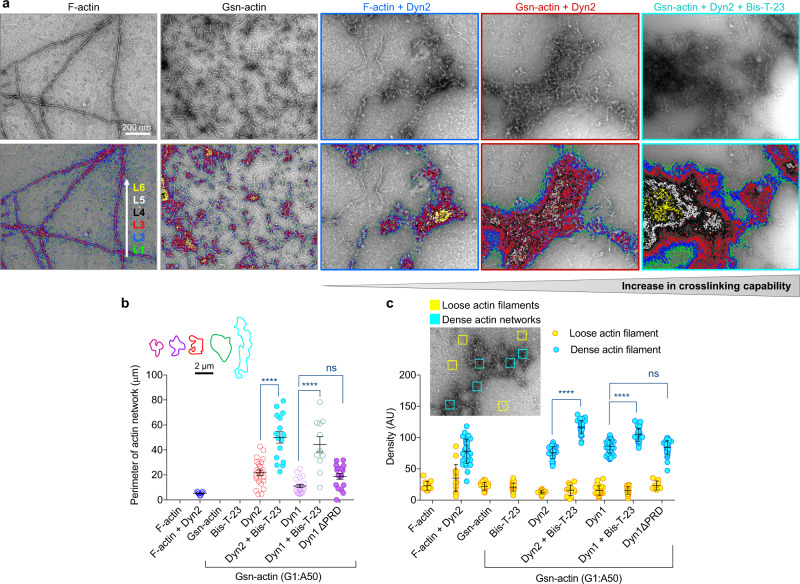
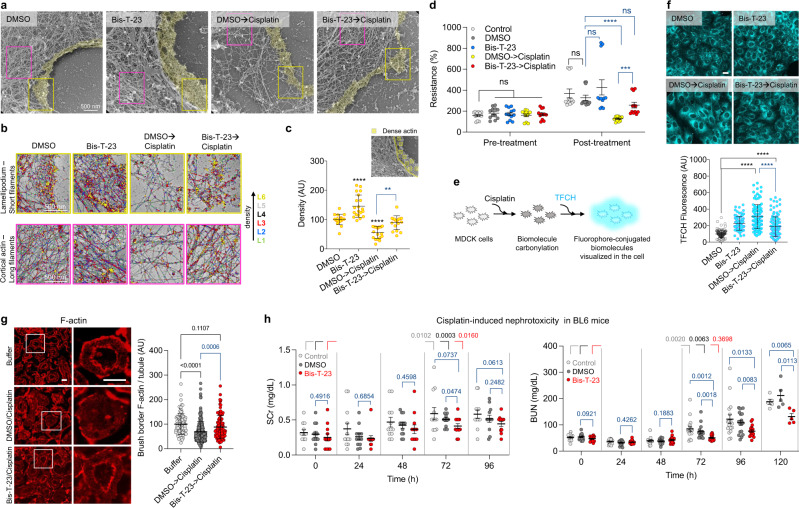
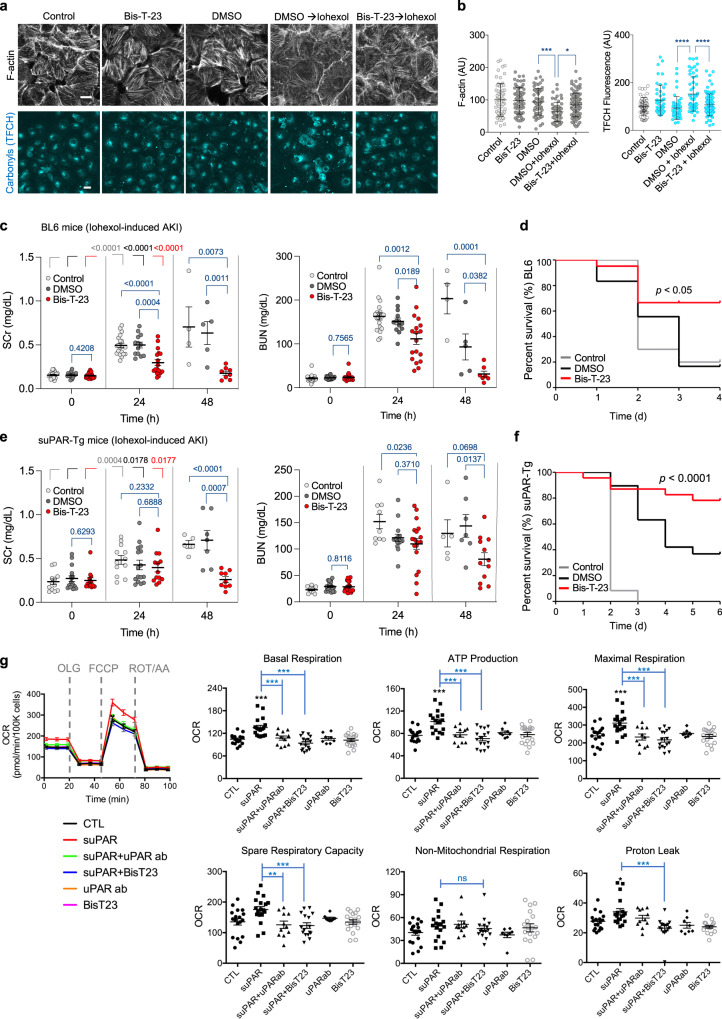
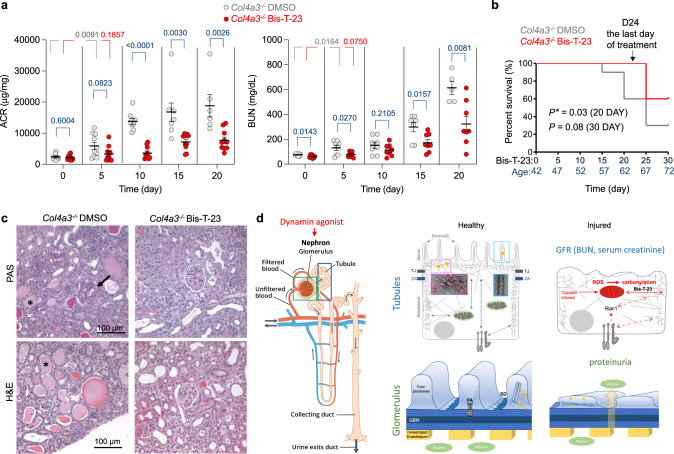
Chronic kidney diseases and acute kidney injury are mechanistically distinct kidney diseases. While chronic kidney diseases are associated with podocyte injury, acute kidney injury affects renal tubular epithelial cells. Despite these differences, a cardinal feature of both acute and chronic kidney diseases is dysregulated actin cytoskeleton. We have shown that pharmacological activation of GTPase dynamin ameliorates podocyte injury in murine models of chronic kidney diseases by promoting actin polymerization. Here we establish dynamin's role in modulating stiffness and polarity of renal tubular epithelial cells by crosslinking actin filaments into branched networks. Activation of dynamin's crosslinking capability by a small molecule agonist stabilizes the actomyosin cortex of the apical membrane against injury, which in turn preserves renal function in various murine models of acute kidney injury. Notably, a dynamin agonist simultaneously attenuates podocyte and tubular injury in the genetic murine model of Alport syndrome. Our study provides evidence for the feasibility and highlights the benefits of novel holistic nephron-protective therapies.
© 2022. The Author(s).
Conflict of interest statement
S.S. and J.R. are co-founders and shareholders of Walden Biosciences, a biotechnology company that develops novel kidney-protective therapies. S.L.B. and K.M. are inventors on a pending patent application pertaining to the detection of oxidative stress-induced carbonylation. The remaining authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
