

New insights into the molecular mechanisms of ROR1, ROR2, and PTK7 signaling from the proteomics and pharmacological modulation of ROR1 interactome
- PMID: 35504983
- PMCID: PMC9064840
- DOI: 10.1007/s00018-022-04301-6
New insights into the molecular mechanisms of ROR1, ROR2, and PTK7 signaling from the proteomics and pharmacological modulation of ROR1 interactome
Abstract
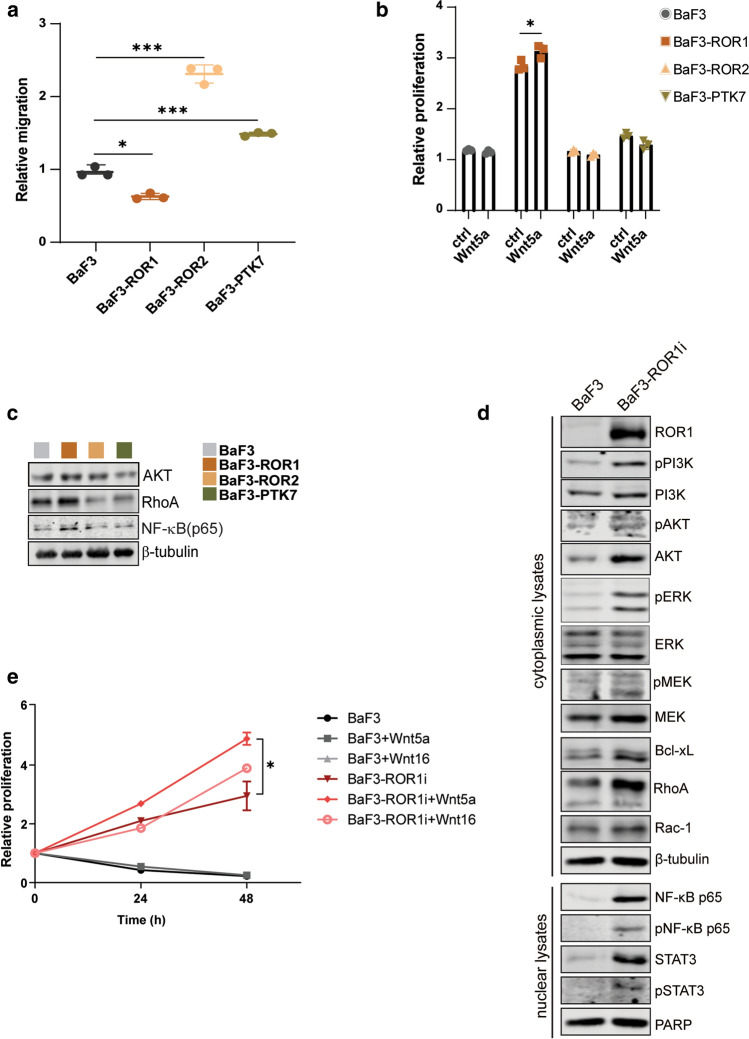
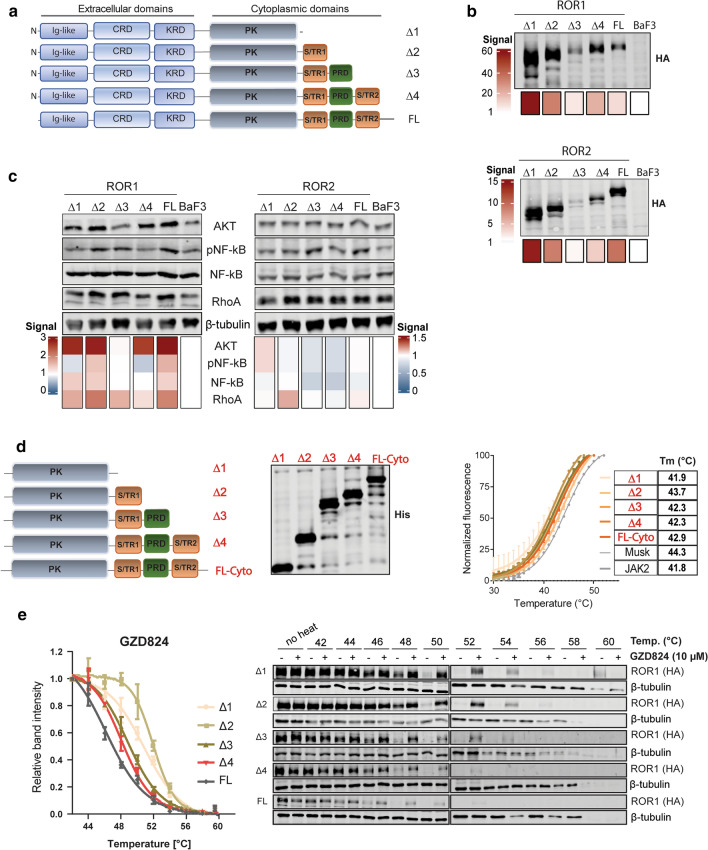
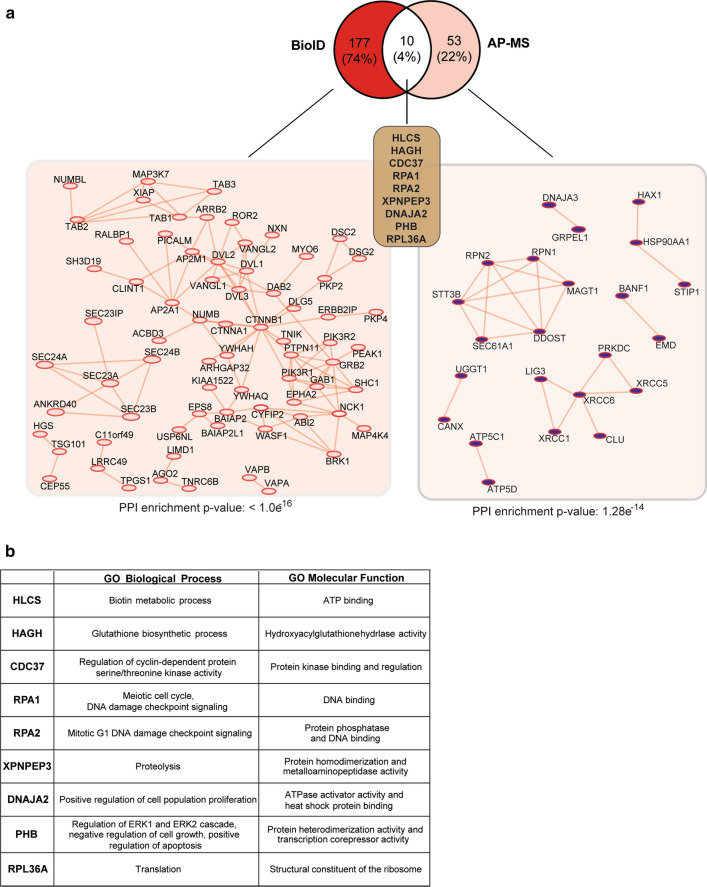
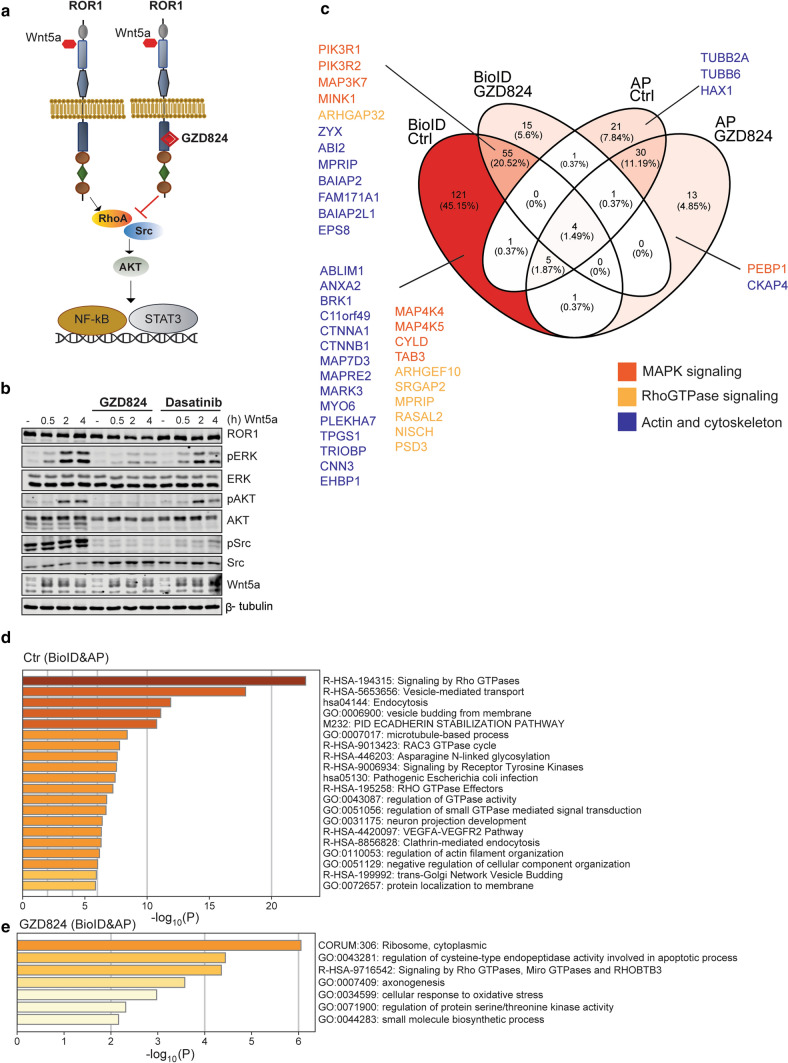
ROR1, ROR2, and PTK7 are Wnt ligand-binding members of the receptor tyrosine kinase family. Despite their lack of catalytic activity, these receptors regulate skeletal, cardiorespiratory, and neurological development during embryonic and fetal stages. However, their overexpression in adult tissue is strongly connected to tumor development and metastasis, suggesting a strong pharmacological potential for these molecules. Wnt5a ligand can activate these receptors, but lead to divergent signaling and functional outcomes through mechanisms that remain largely unknown. Here, we developed a cellular model by stably expressing ROR1, ROR2, and PTK7 in BaF3 cells that allowed us to readily investigate side-by-side their signaling capability and functional outcome. We applied proteomic profiling to BaF3 clones and identified distinctive roles for ROR1, ROR2, and PTK7 pseudokinases in modulating the expression of proteins involved in cytoskeleton dynamics, apoptotic, and metabolic signaling. Functionally, we show that ROR1 expression enhances cell survival and Wnt-mediated cell proliferation, while ROR2 and PTK7 expression is linked to cell migration. We also demonstrate that the distal C-terminal regions of ROR1 and ROR2 are required for receptors stability and downstream signaling. To probe the pharmacological modulation of ROR1 oncogenic signaling, we used affinity purification coupled to mass spectrometry (AP-MS) and proximity-dependent biotin identification (BioID) to map its interactome before and after binding of GZD824, a small molecule inhibitor previously shown to bind to the ROR1 pseudokinase domain. Our findings bring new insight into the molecular mechanisms of ROR1, ROR2, and PTK7, and highlight the therapeutic potential of targeting ROR1 with small molecule inhibitors binding to its vestigial ATP-binding site.
Keywords: CETSA; GZD824; Kinase inhibitors; PTK7; ROR1; ROR2; Receptor pseudokinases; TSA; Wnt signaling.
© 2022. The Author(s).
Conflict of interest statement
The authors have no relevant financial or non-financial interests to disclose.
Figures

References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
