

Tbx2 is a master regulator of inner versus outer hair cell differentiation
- PMID: 35508658
- PMCID: PMC9803360
- DOI: 10.1038/s41586-022-04668-3
Tbx2 is a master regulator of inner versus outer hair cell differentiation
Abstract
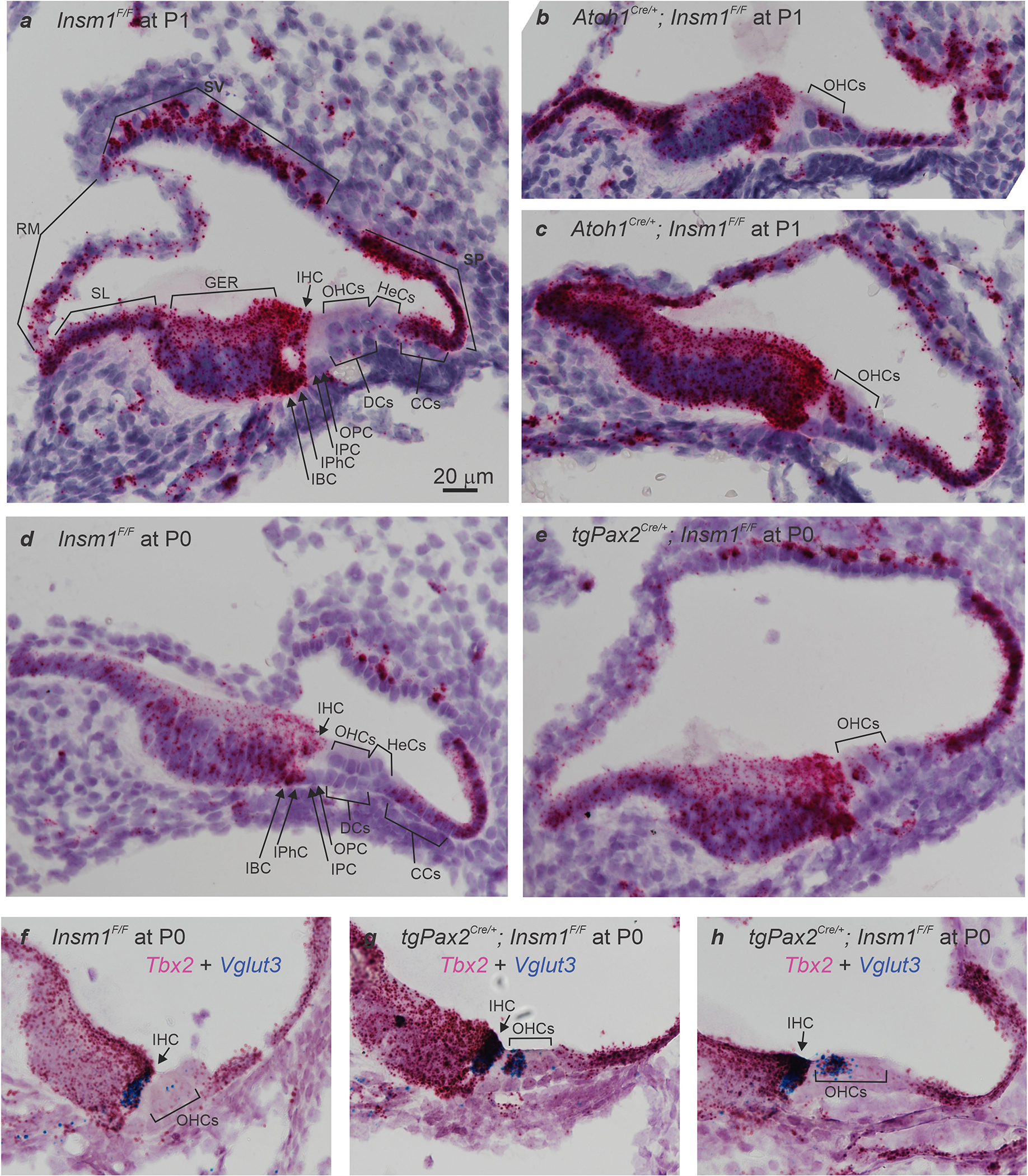
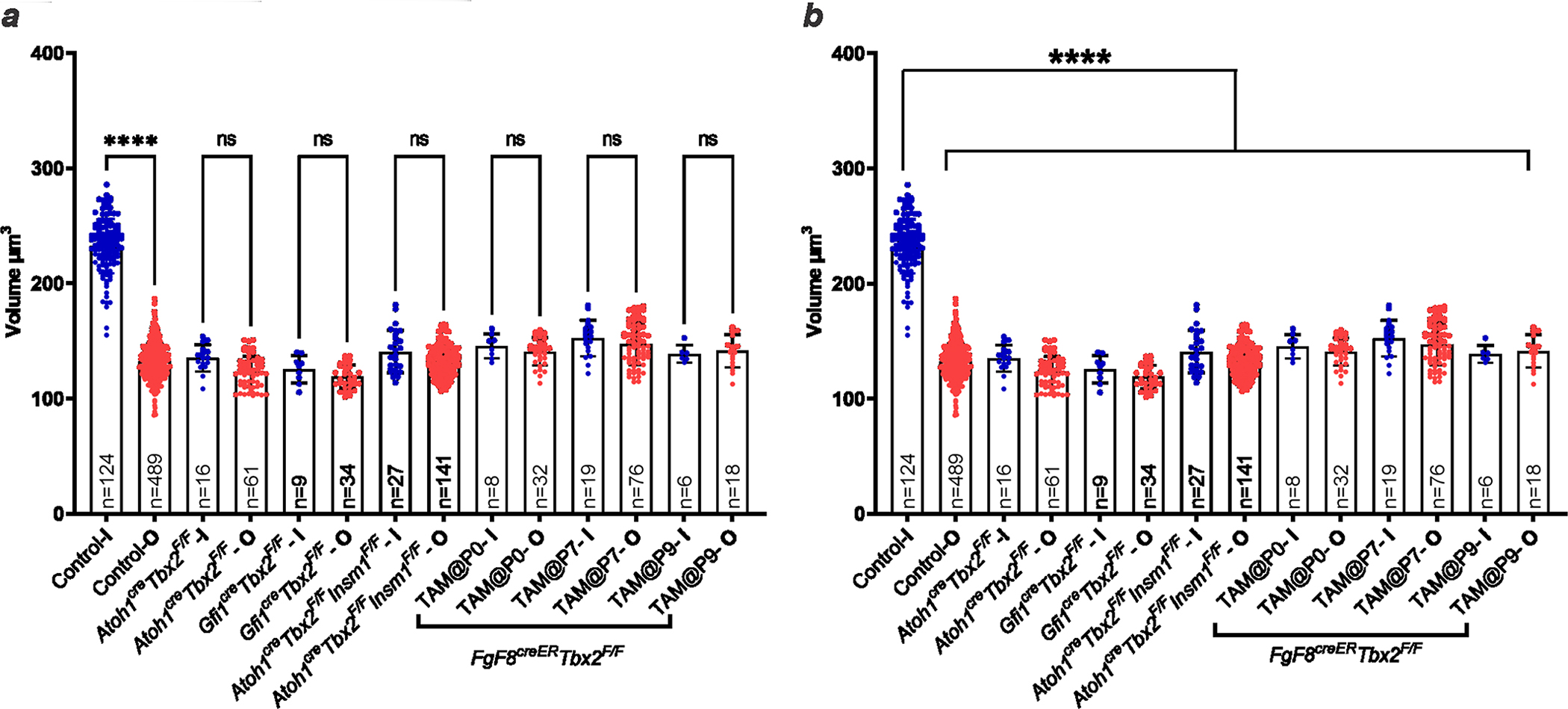
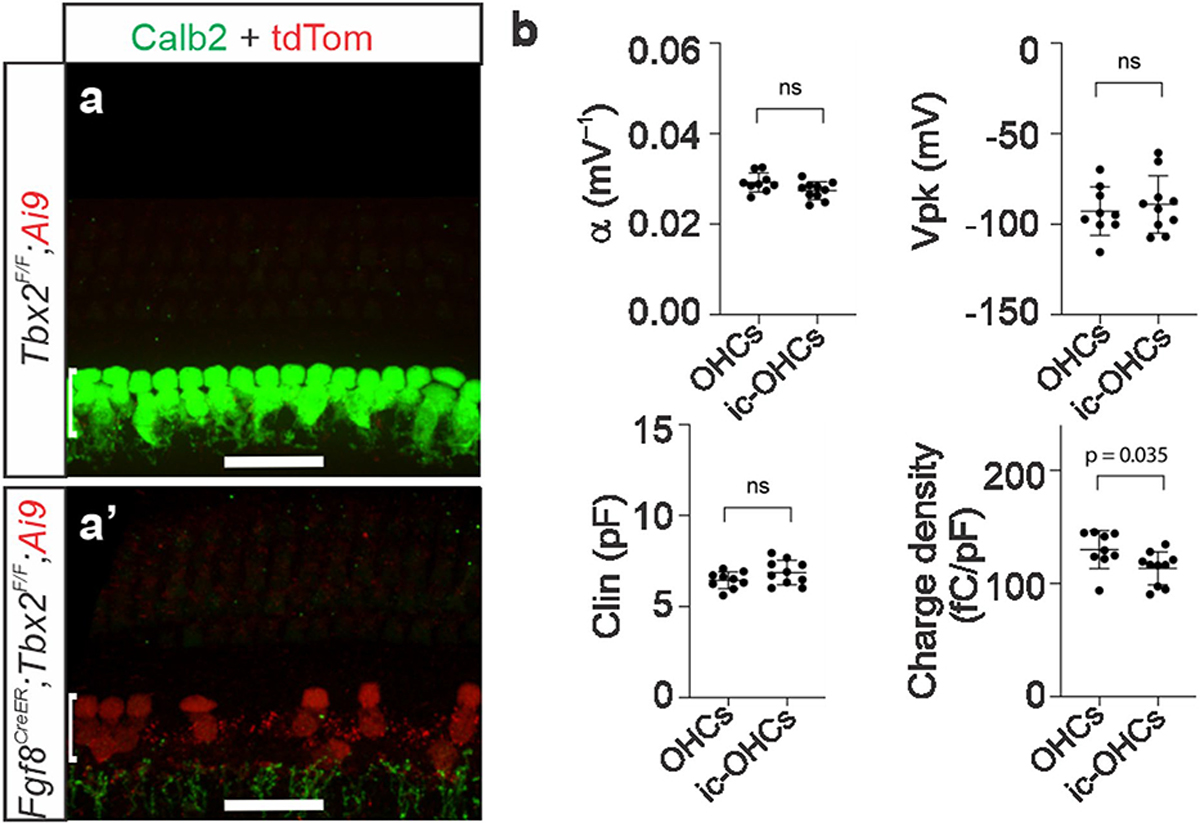
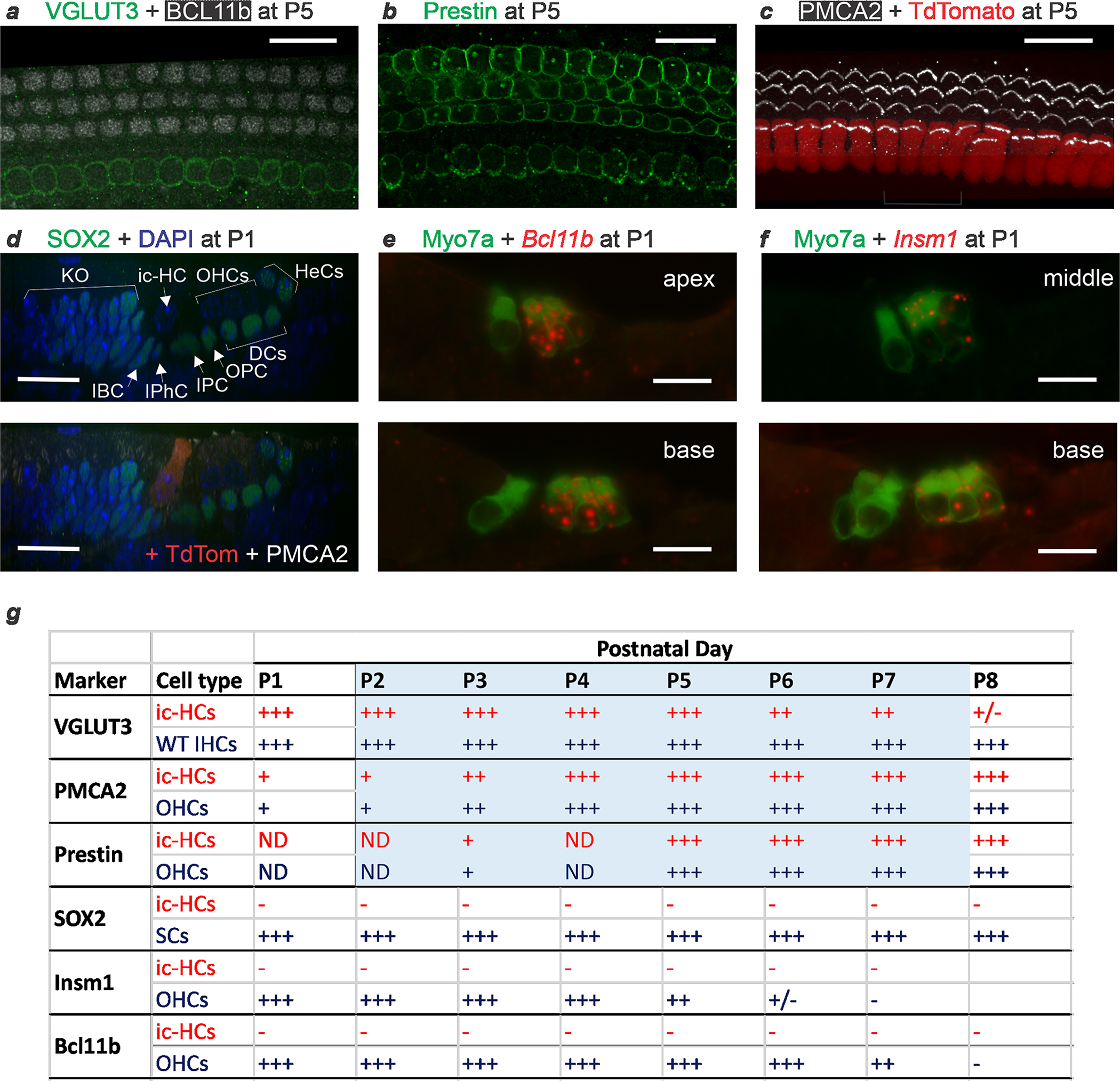
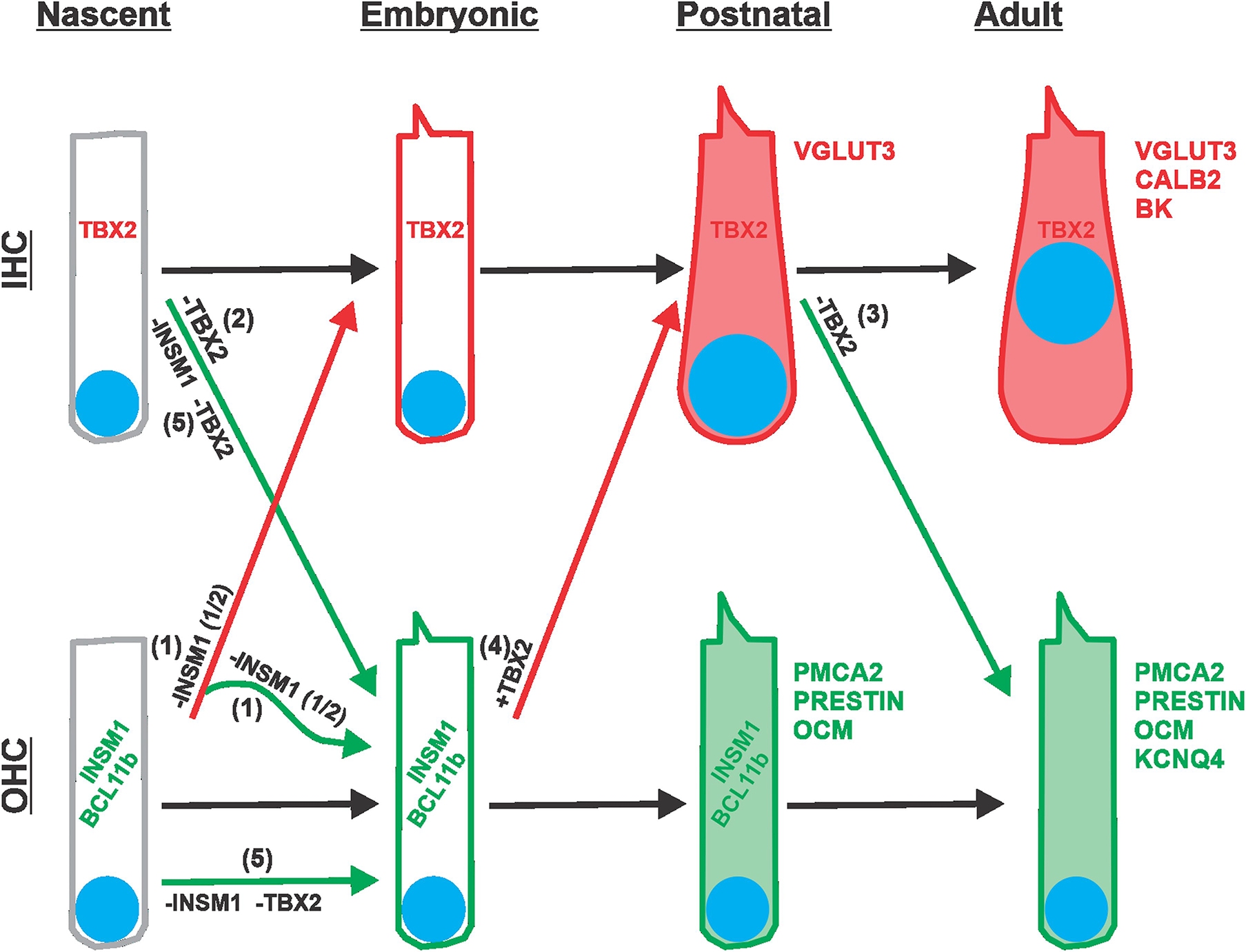
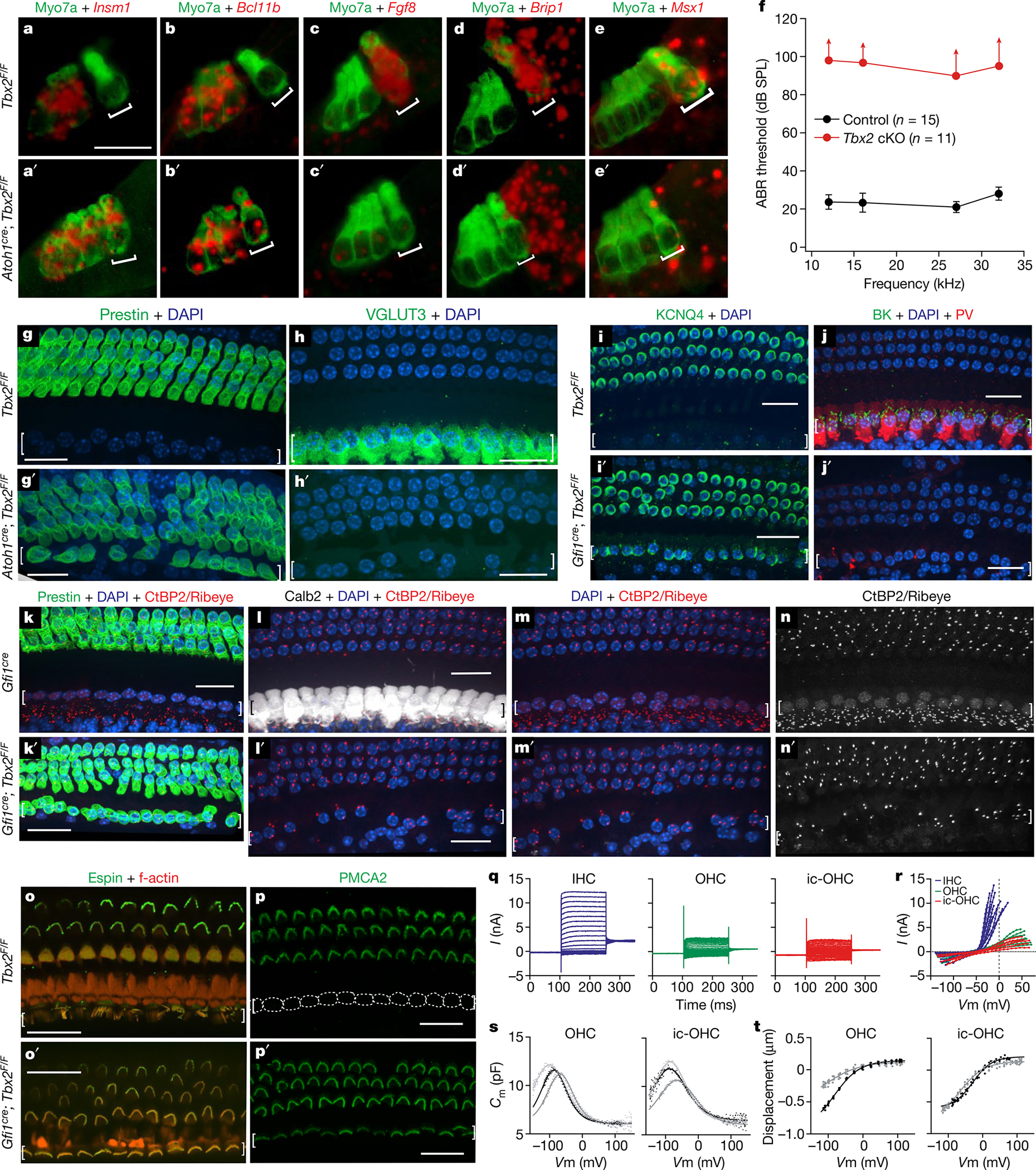
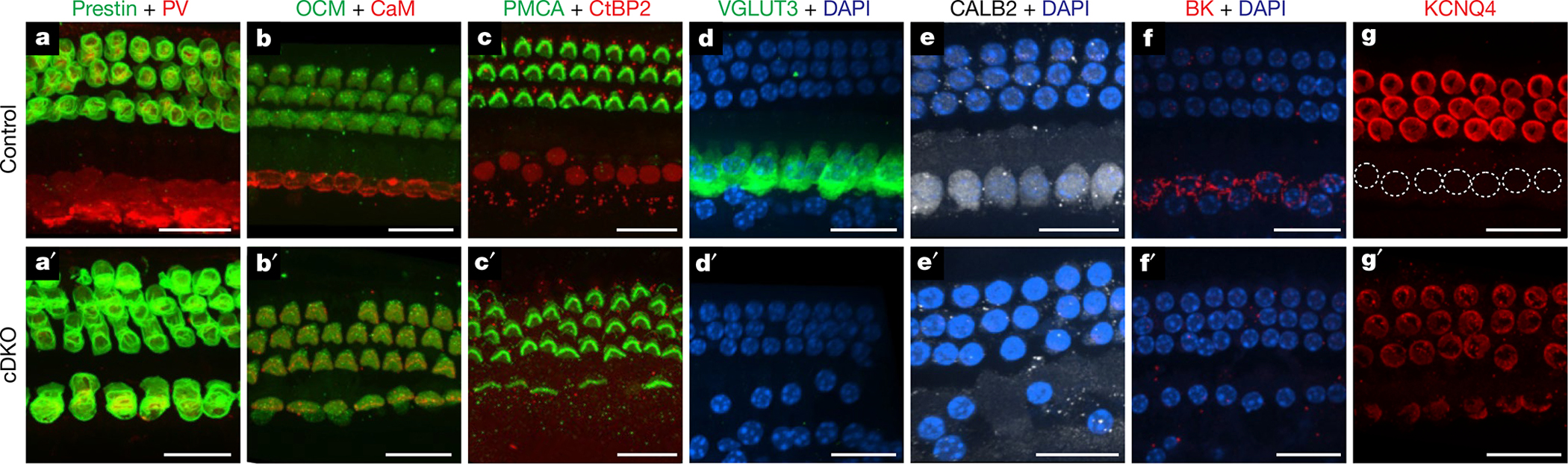
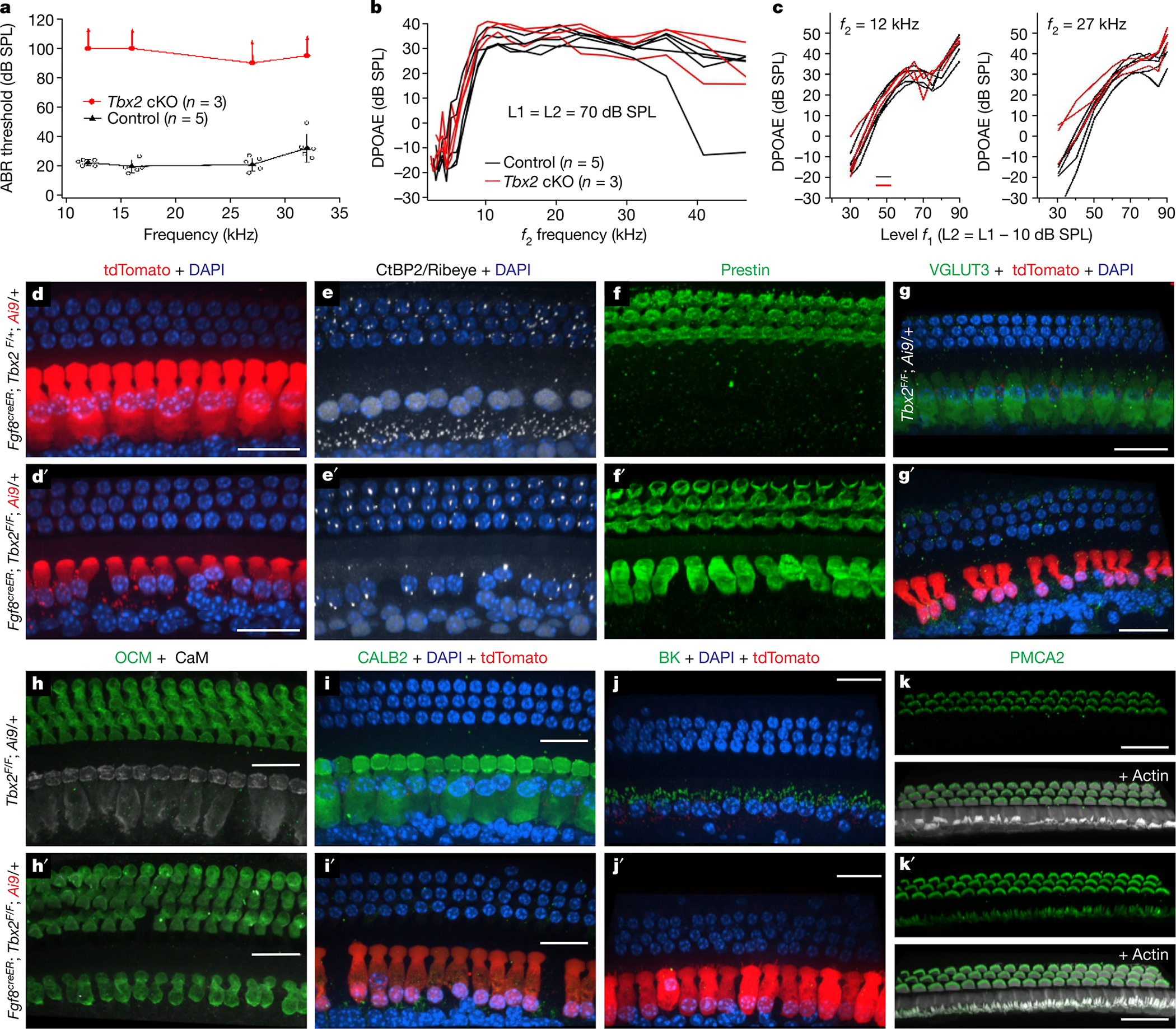
The cochlea uses two types of mechanosensory cell to detect sounds. A single row of inner hair cells (IHCs) synapse onto neurons to transmit sensory information to the brain, and three rows of outer hair cells (OHCs) selectively amplify auditory inputs1. So far, two transcription factors have been implicated in the specific differentiation of OHCs, whereas, to our knowledge, none has been identified in the differentiation of IHCs2-4. One such transcription factor for OHCs, INSM1, acts during a crucial embryonic period to consolidate the OHC fate, preventing OHCs from transdifferentiating into IHCs2. In the absence of INSM1, embryonic OHCs misexpress a core set of IHC-specific genes, which we predict are involved in IHC differentiation. Here we find that one of these genes, Tbx2, is a master regulator of IHC versus OHC differentiation in mice. Ablation of Tbx2 in embryonic IHCs results in their development as OHCs, expressing early OHC markers such as Insm1 and eventually becoming completely mature OHCs in the position of IHCs. Furthermore, Tbx2 is epistatic to Insm1: in the absence of both genes, cochleae generate only OHCs, which suggests that TBX2 is necessary for the abnormal transdifferentiation of INSM1-deficient OHCs into IHCs, as well as for normal IHC differentiation. Ablation of Tbx2 in postnatal, largely differentiated IHCs makes them transdifferentiate directly into OHCs, replacing IHC features with those of mature and not embryonic OHCs. Finally, ectopic expression of Tbx2 in OHCs results in their transdifferentiation into IHCs. Hence, Tbx2 is both necessary and sufficient to make IHCs distinct from OHCs and maintain this difference throughout development.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
References
-
- Dallos P In Proceedings of the 9th International Symposium on Hearing (eds Cazals Y et al.) 3–17 (Elsevier, 1992).
-
- Kaiser M et al. Regulation of otocyst patterning by Tbx2 and Tbx3 is required for inner ear morphogenesis in the mouse. Development 148, dev195651 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
