

The degree of unsaturation of fatty acids in phosphatidylserine alters the rate of insulin aggregation and the structure and toxicity of amyloid aggregates
- PMID: 35510803
- PMCID: PMC9197964
- DOI: 10.1002/1873-3468.14369
The degree of unsaturation of fatty acids in phosphatidylserine alters the rate of insulin aggregation and the structure and toxicity of amyloid aggregates
Abstract
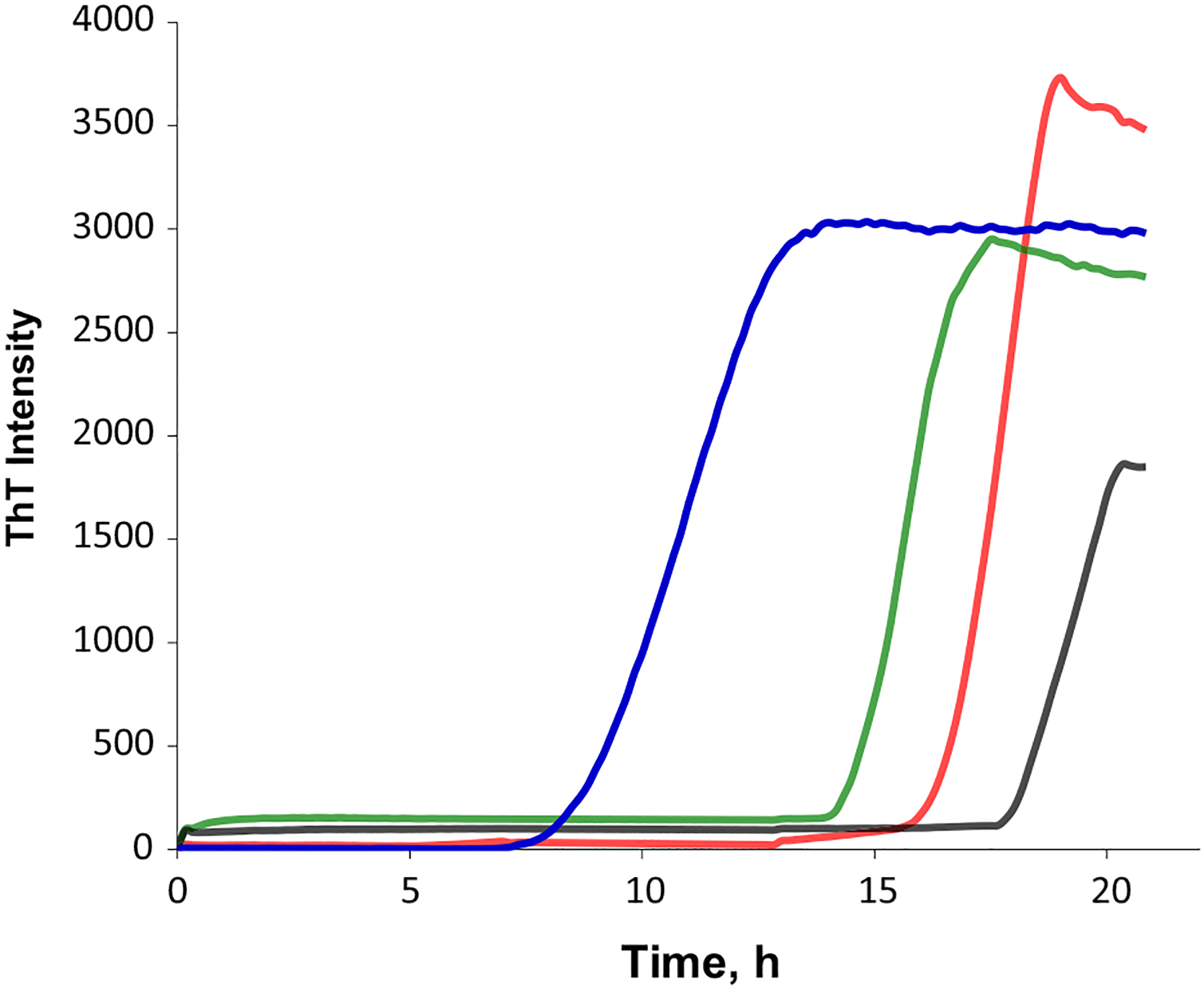
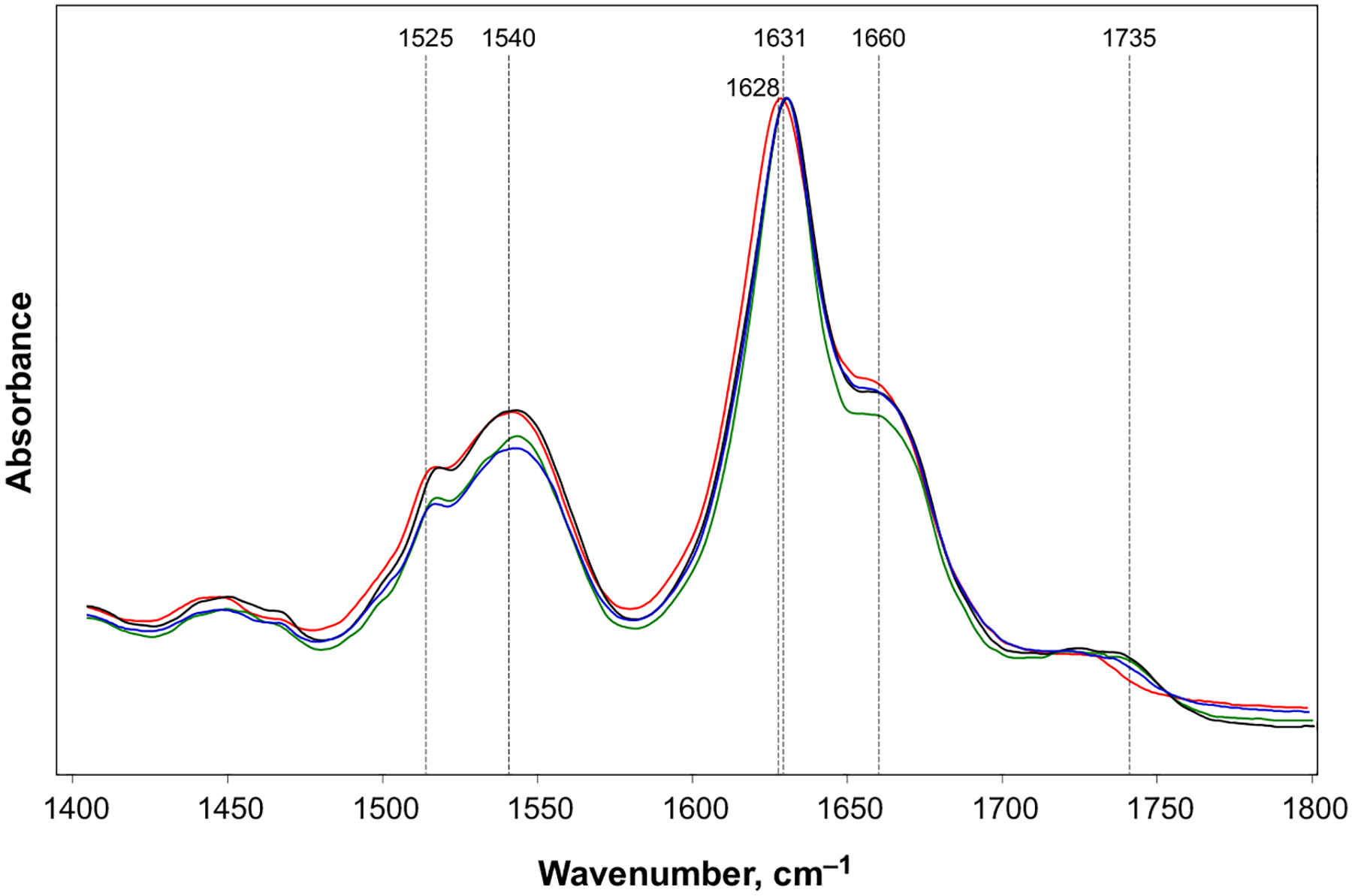
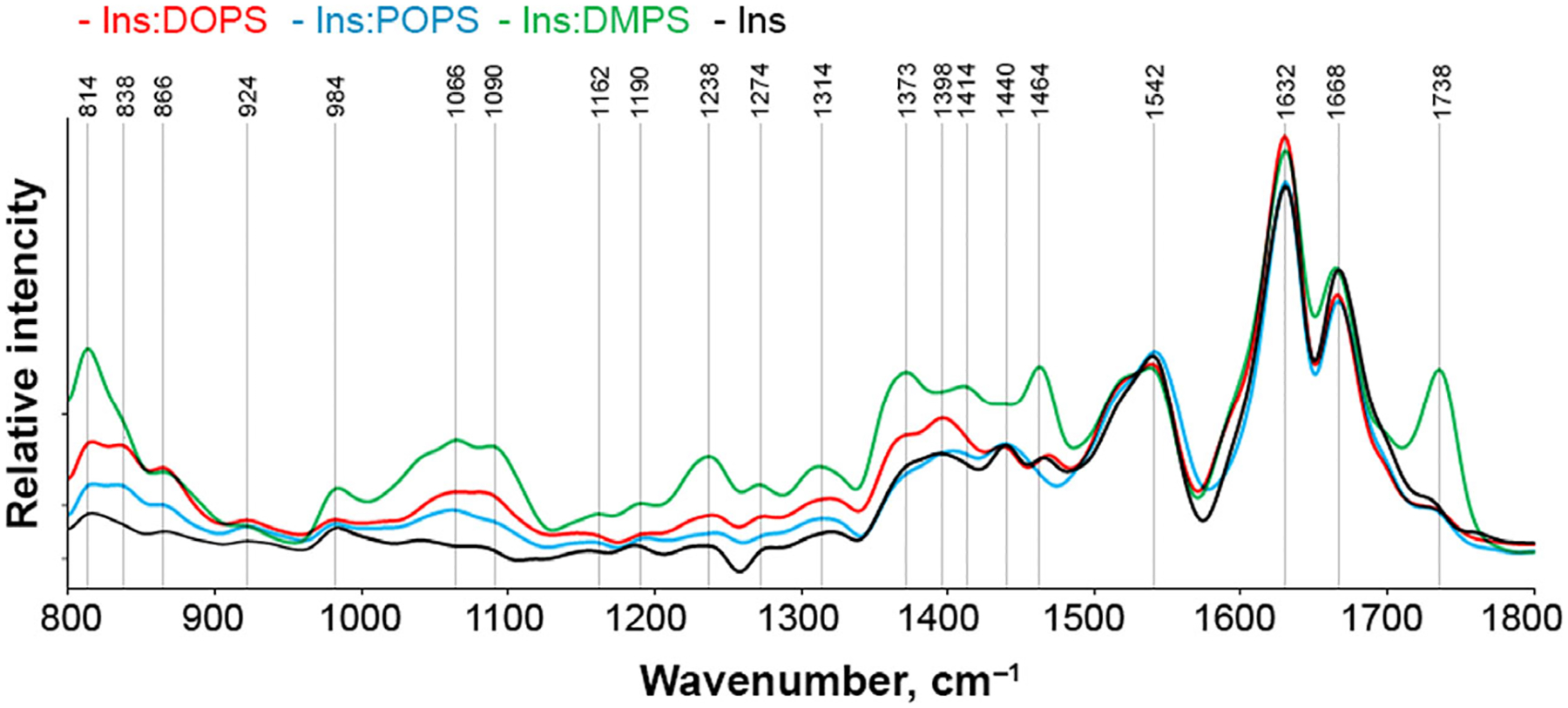
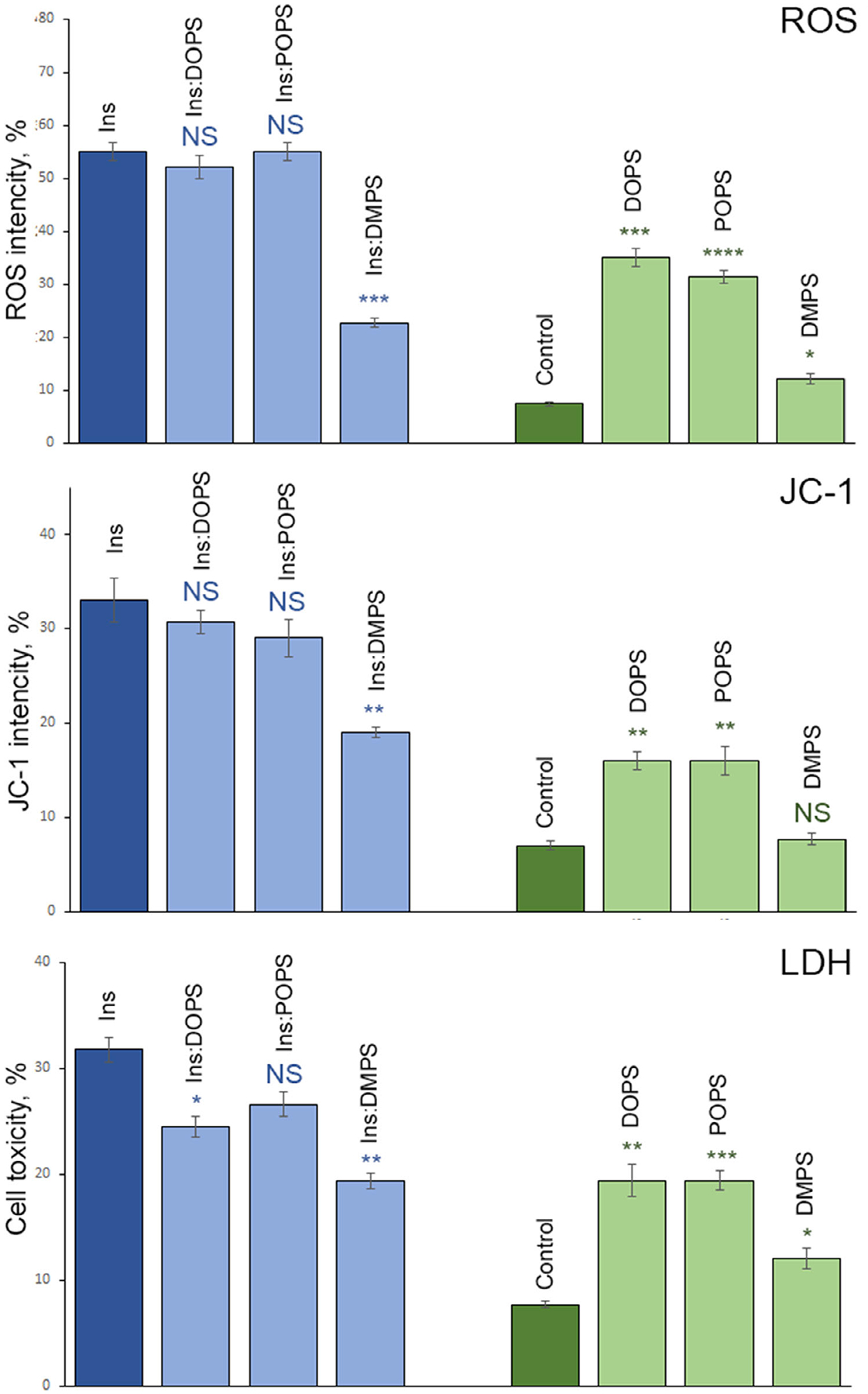
Phosphatidylserine (PS) in the plasma membrane plays an important role in cell signaling and apoptosis. Cell degeneration is also linked to numerous amyloid diseases, pathologies that are associated with aggregation of misfolded proteins. In this work, we examine the effect of both saturated PS (DMPS) and unsaturated PS (DOPS and POPS) on the aggregation properties of insulin, as well as the structure and toxicity of insulin aggregates formed in the presence of these phospholipids. We found that the degree of unsaturation of fatty acids in PS alters the rate of insulin aggregation. We also found that toxicity of insulin-DMPS aggregates is significantly lower than the toxicity of DOPS- and POPS-insulin fibrils, whereas all these lipid-containing aggregates exert lower cell toxicity than insulin fibrils grown in a lipid-free environment.
Keywords: AFM-IR; amyloid; insulin; phospholipids; toxicity.
© 2022 Federation of European Biochemical Societies.
Figures

References
-
- Chiti F, Dobson CM. Protein misfolding, amyloid formation, and human disease: a summary of progress over the last decade. Annu Rev Biochem. 2017;86:27–68. - PubMed
-
- Knowles TP, Vendruscolo M, Dobson CM. The amyloid state and its association with protein misfolding diseases. Nat Rev Mol Cell Biol. 2014;15:384–96. - PubMed
-
- Dou T, Zhou L, Kurouski D. Unravelling the structural organization of individual alpha-synuclein oligomers grown in the presence of phospholipids. J Phys Chem Lett. 2021;12:4407–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
