

Molecular analysis and essentiality of Aro1 shikimate biosynthesis multi-enzyme in Candida albicans
- PMID: 35512834
- PMCID: PMC9074039
- DOI: 10.26508/lsa.202101358
Molecular analysis and essentiality of Aro1 shikimate biosynthesis multi-enzyme in Candida albicans
Abstract
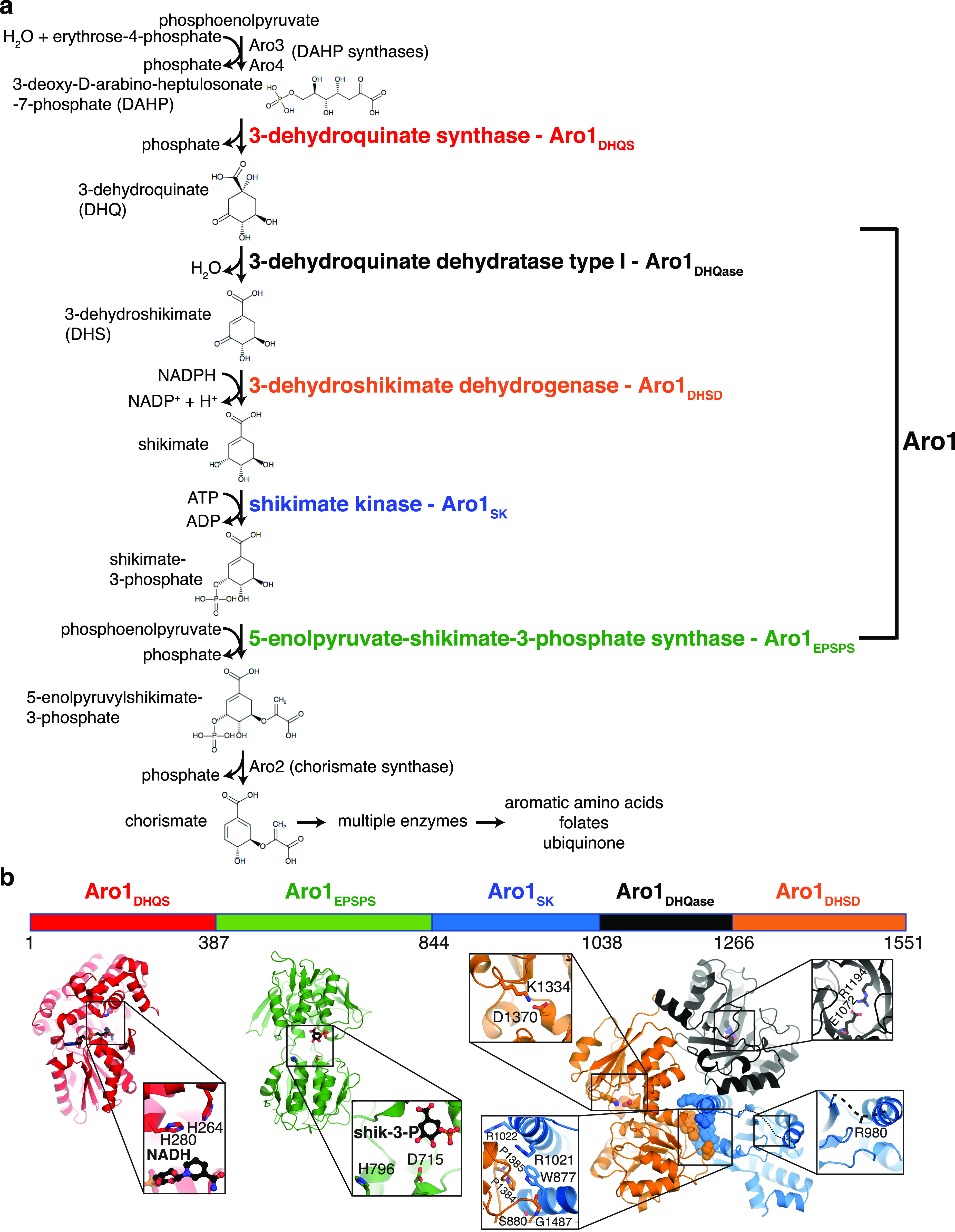
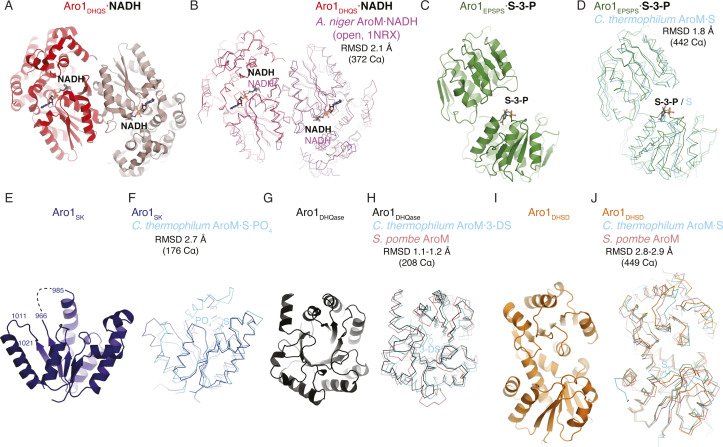
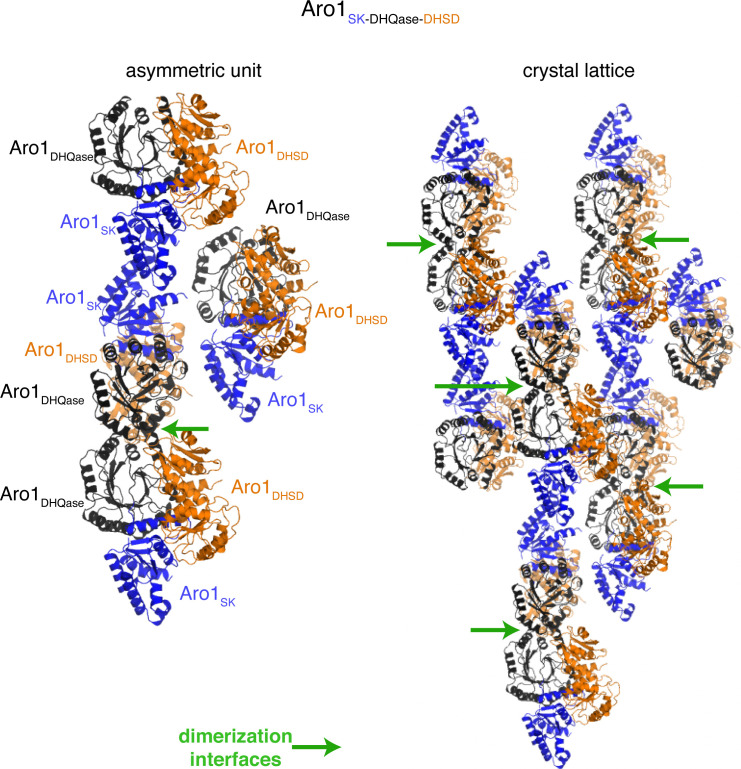
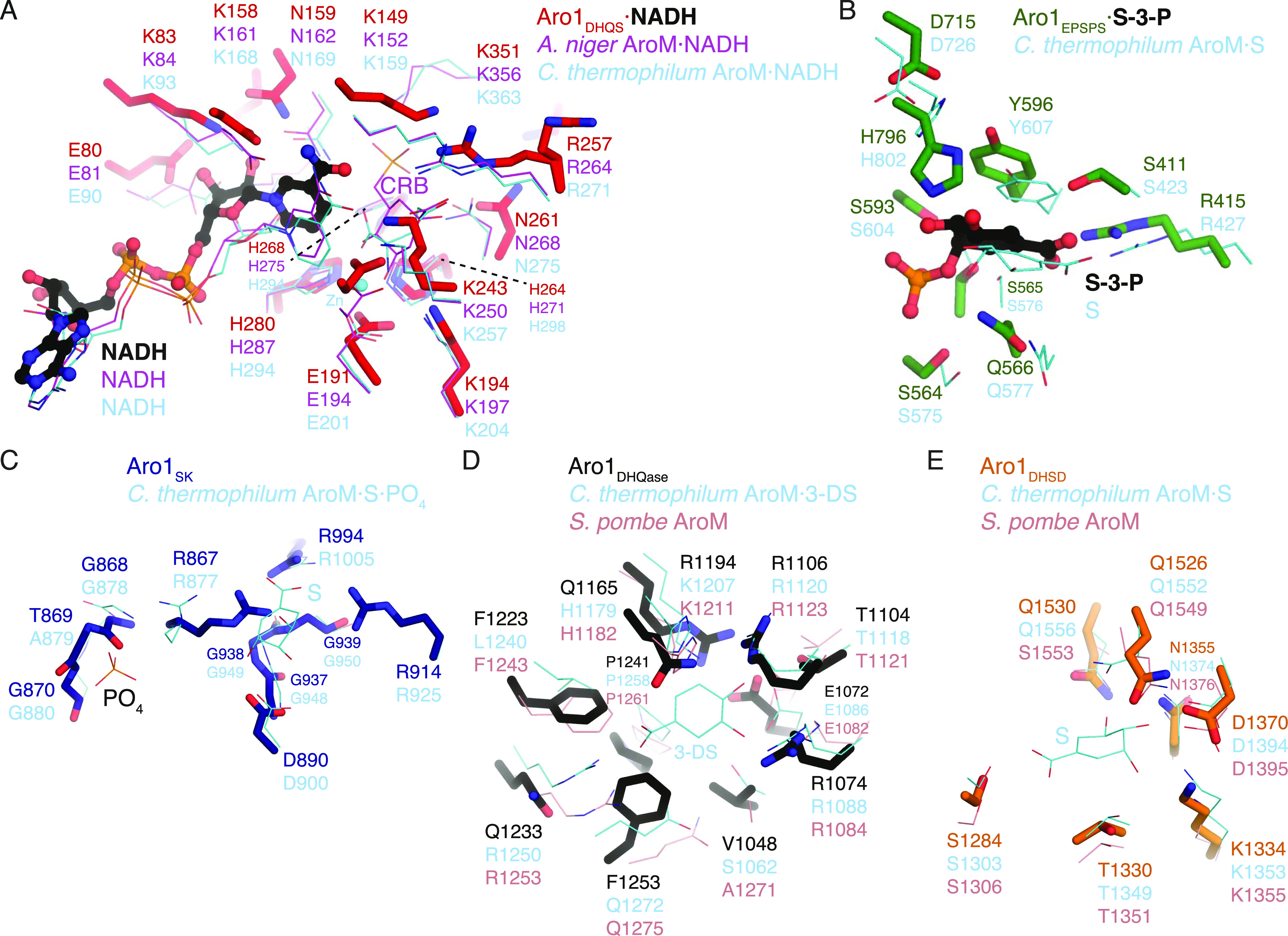
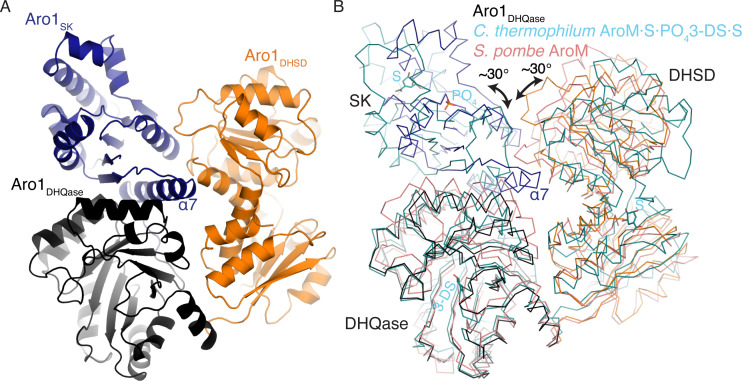
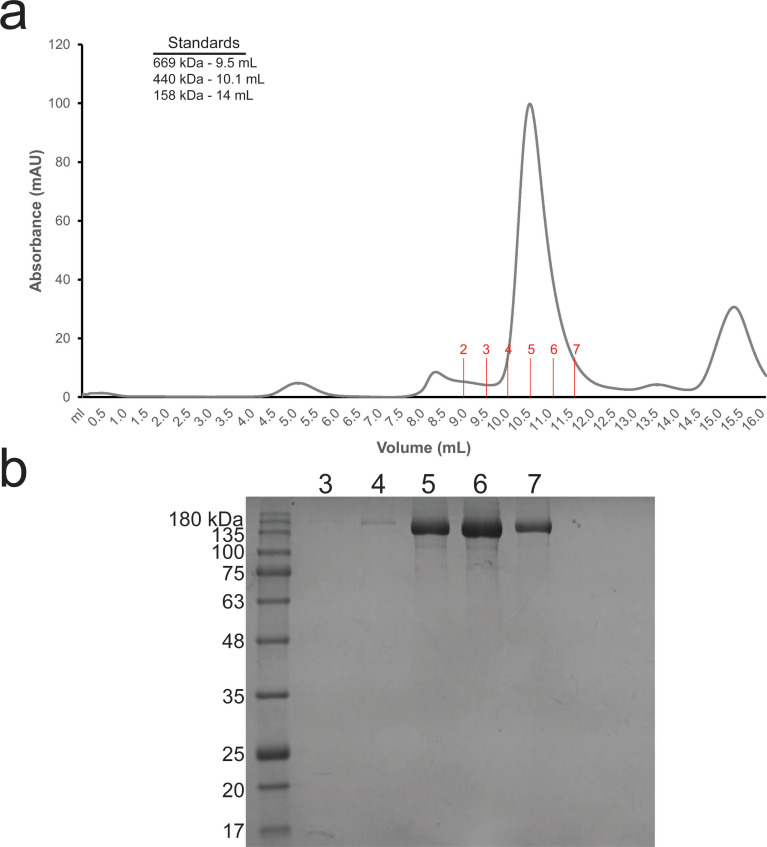
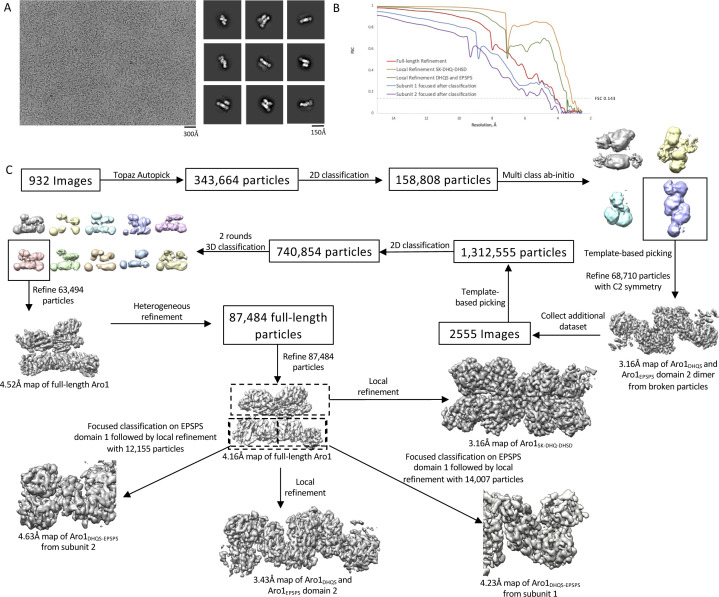
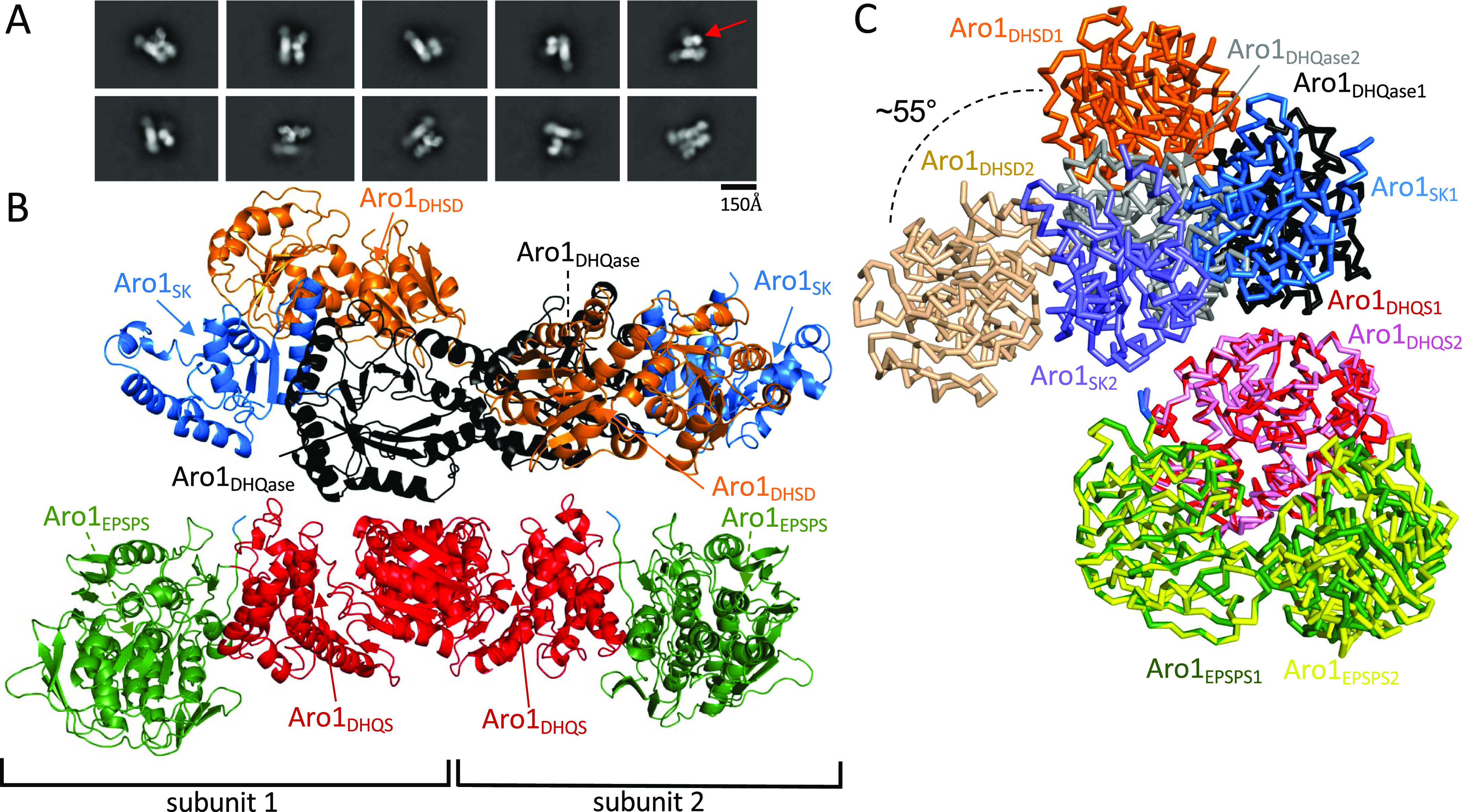
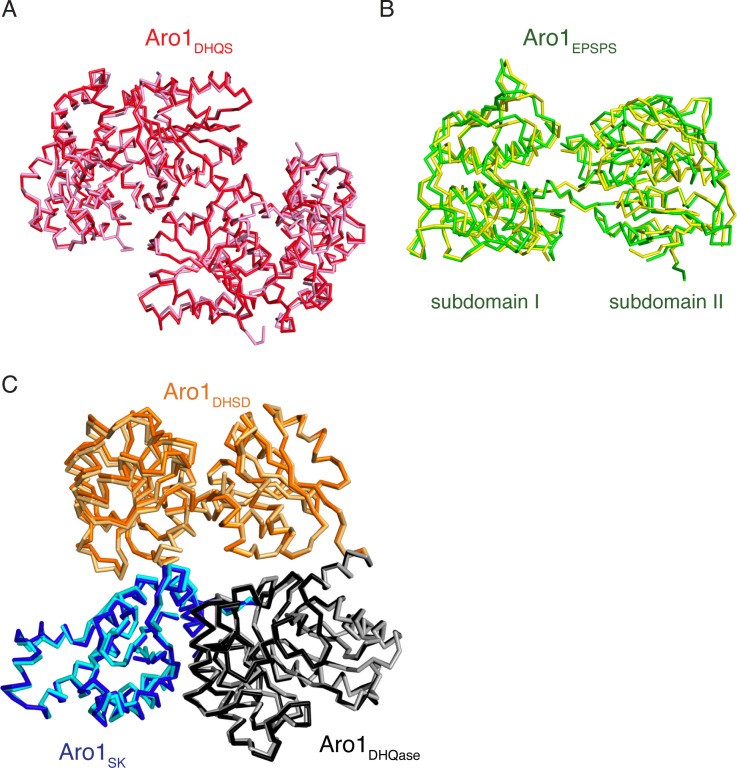
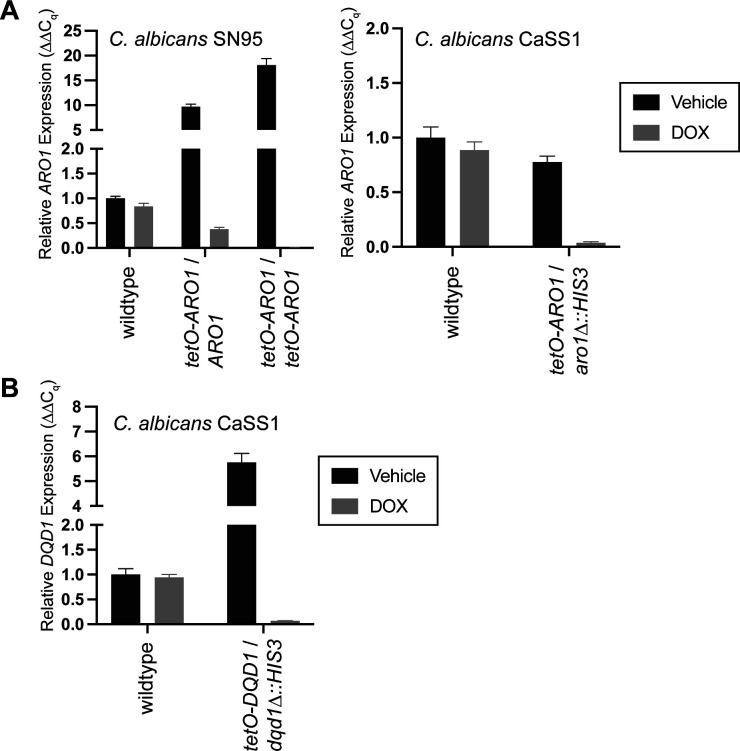
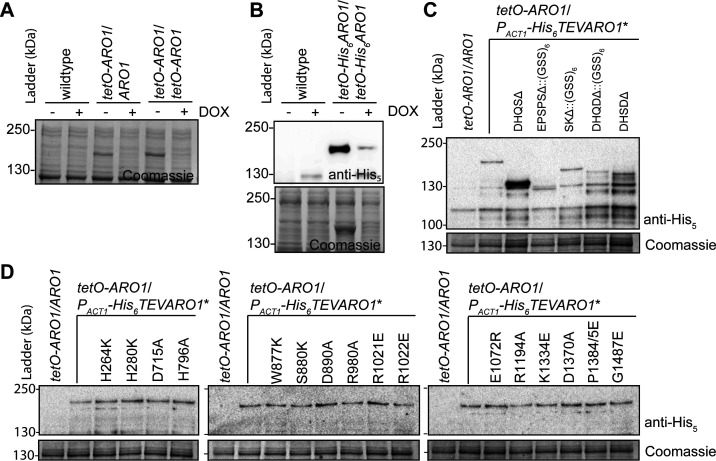
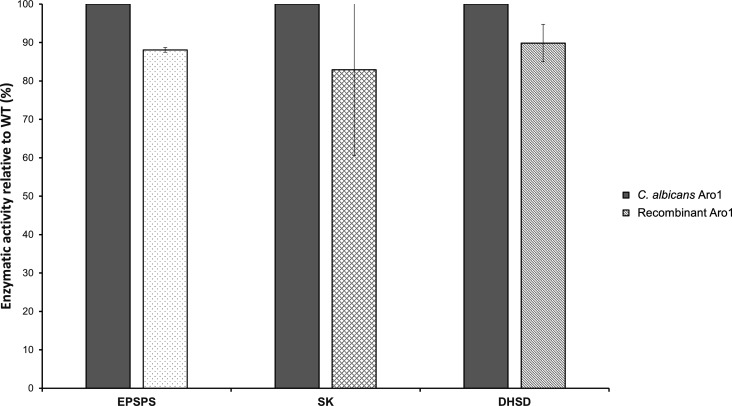
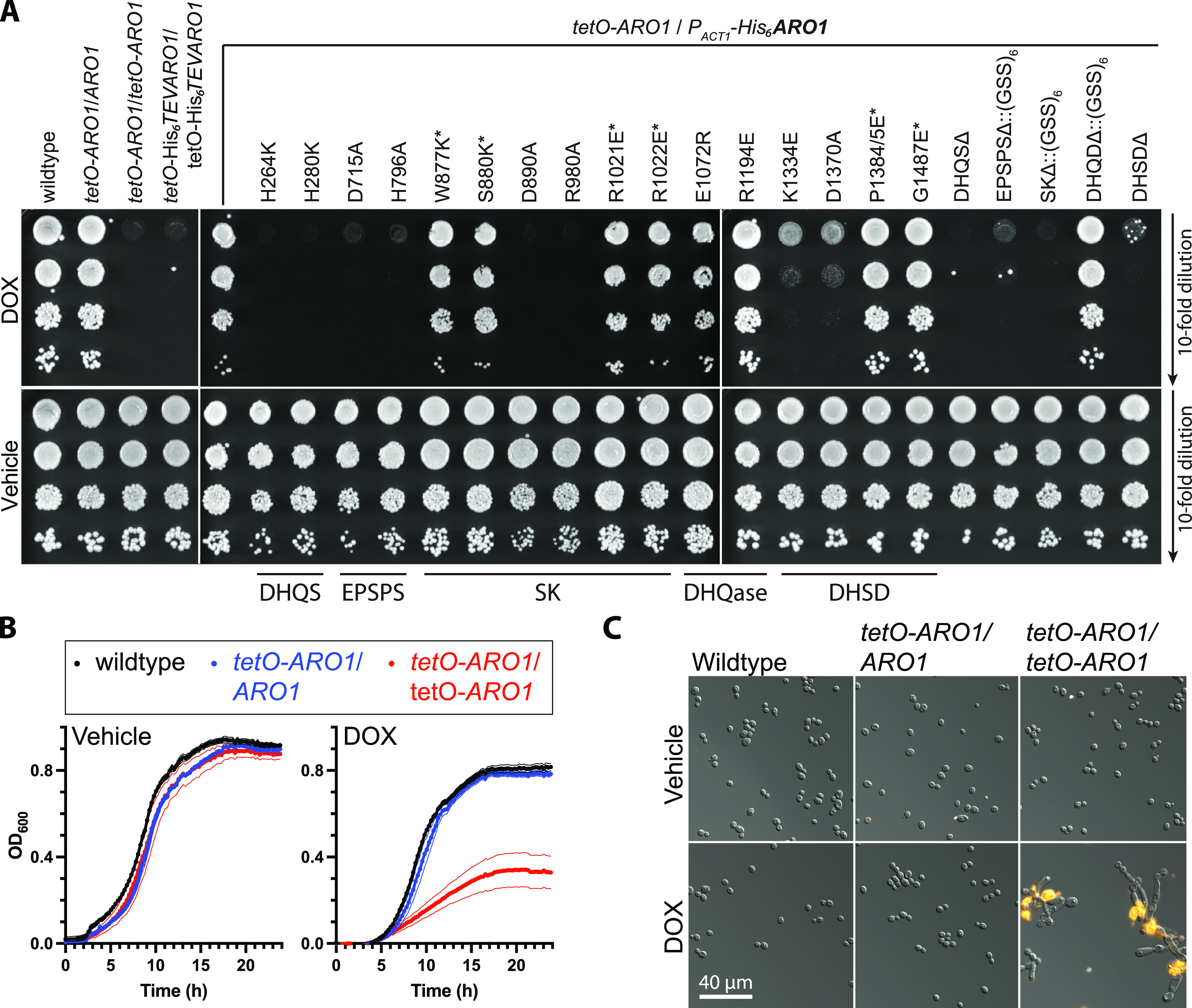
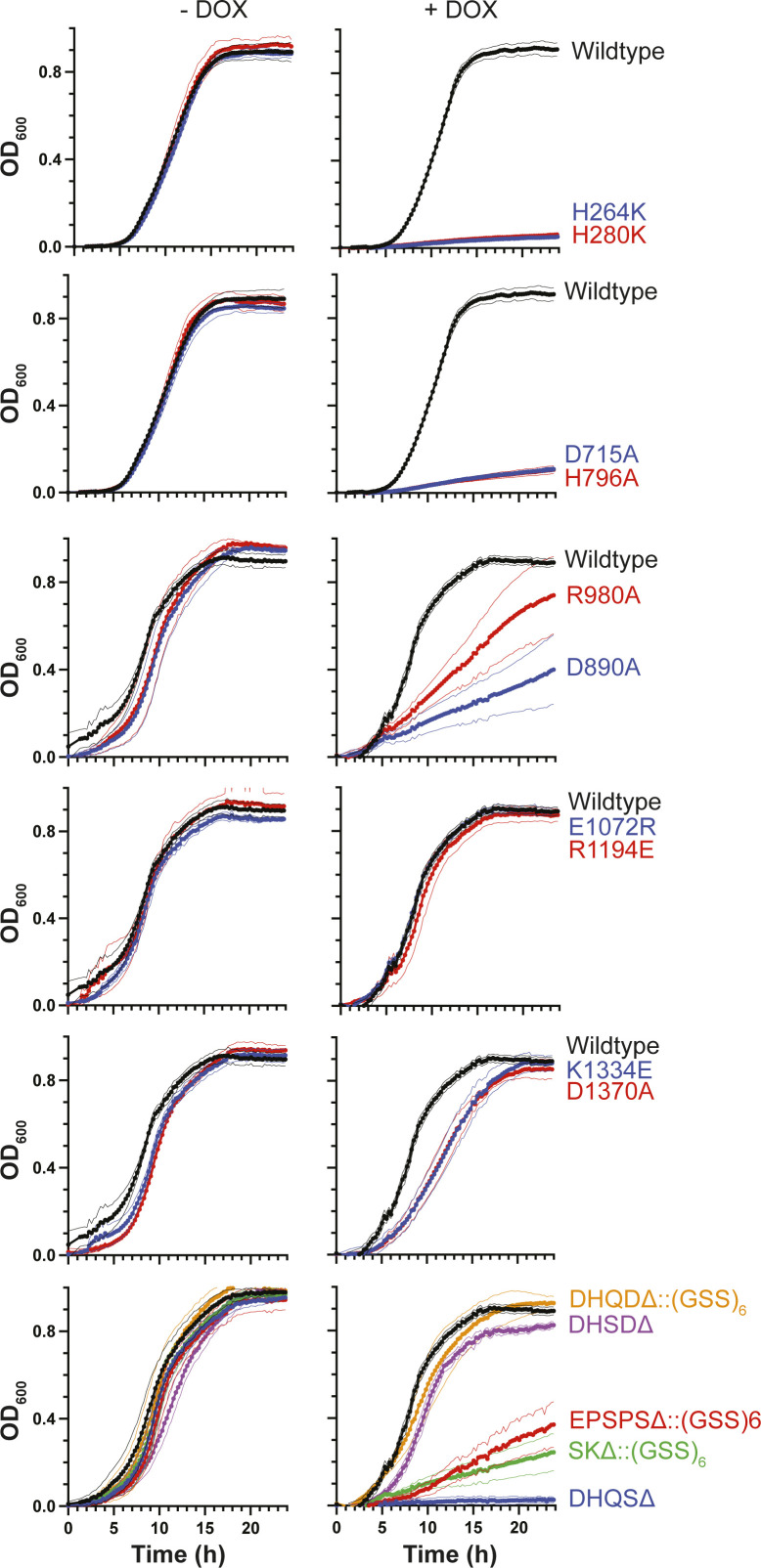
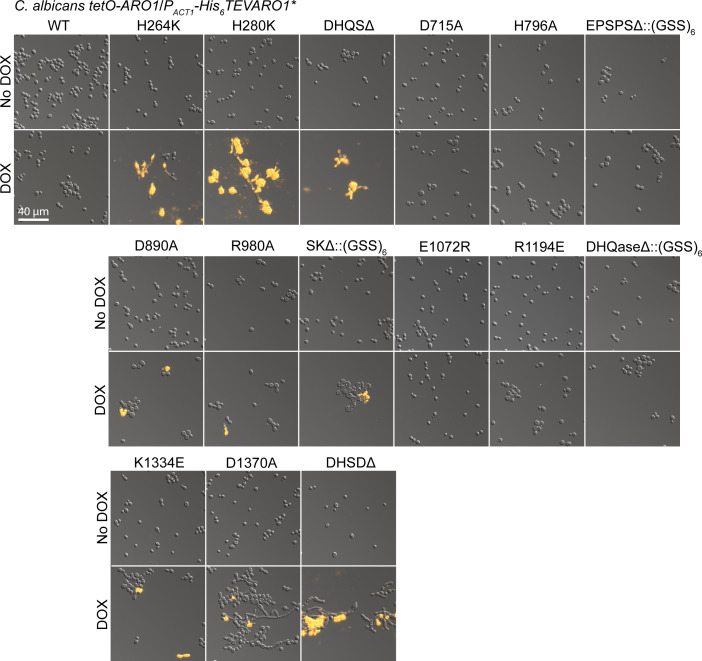
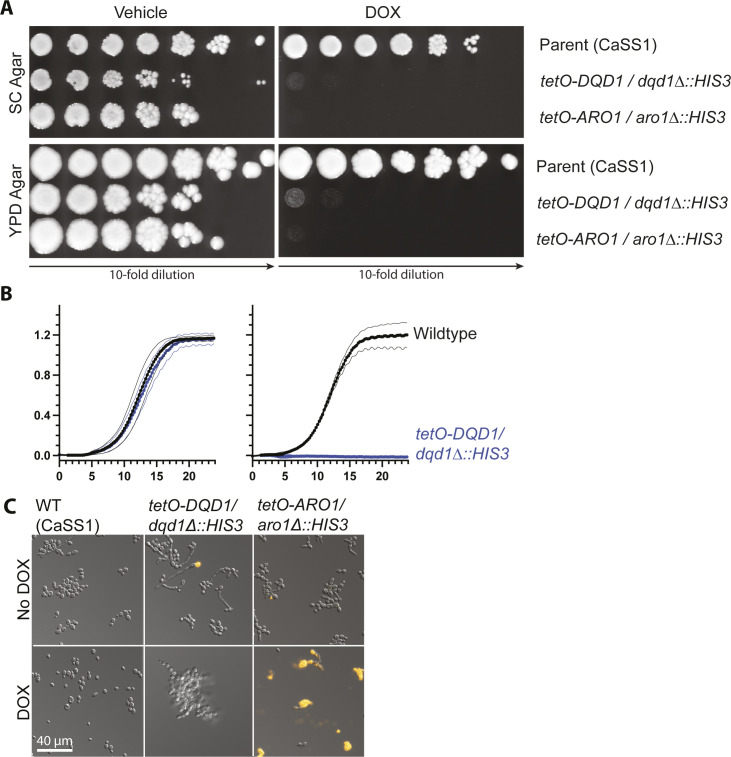
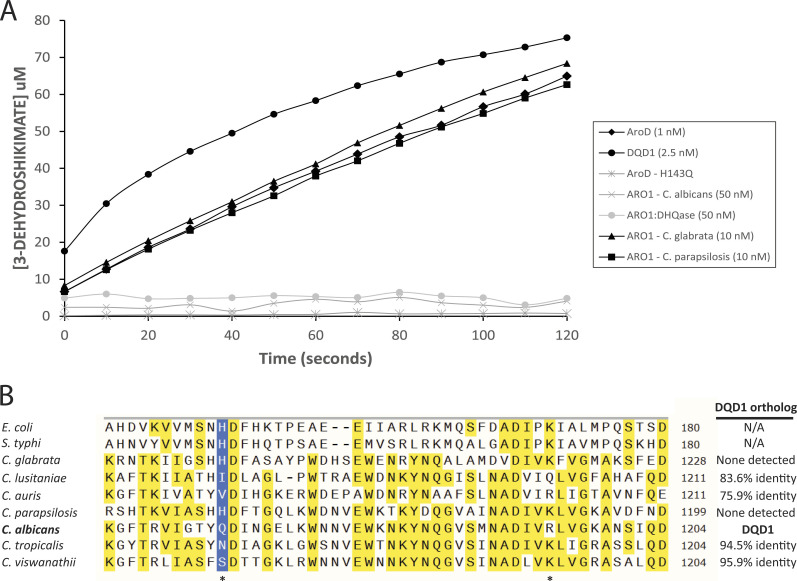
In the human fungal pathogen Candida albicans, ARO1 encodes an essential multi-enzyme that catalyses consecutive steps in the shikimate pathway for biosynthesis of chorismate, a precursor to folate and the aromatic amino acids. We obtained the first molecular image of C. albicans Aro1 that reveals the architecture of all five enzymatic domains and their arrangement in the context of the full-length protein. Aro1 forms a flexible dimer allowing relative autonomy of enzymatic function of the individual domains. Our activity and in cellulo data suggest that only four of Aro1's enzymatic domains are functional and essential for viability of C. albicans, whereas the 3-dehydroquinate dehydratase (DHQase) domain is inactive because of active site substitutions. We further demonstrate that in C. albicans, the type II DHQase Dqd1 can compensate for the inactive DHQase domain of Aro1, suggesting an unrecognized essential role for this enzyme in shikimate biosynthesis. In contrast, in Candida glabrata and Candida parapsilosis, which do not encode a Dqd1 homolog, Aro1 DHQase domains are enzymatically active, highlighting diversity across Candida species.
© 2022 Stogios et al.
Conflict of interest statement
LE Cowen is a co-founder and shareholder in Bright Angel Therapeutics, a platform company for development of novel antifungal therapeutics. LE Cowen is a consultant for Boragen, a small-molecule development company focused on leveraging the unique chemical properties of boron chemistry for crop protection and animal health. SD Liston was supported by a Mitacs postdoctoral fellowship funded in partnership with Amplyx Pharmaceuticals Inc., now a subsidiary of Pfizer. D Borek and Z Otwinowski are co-founders of Ligo Analytics, a company that develops a software for cryogenic electron microscopy. Z Otwinowski is a co-founder of HKL Research, a company that develops software for X-ray diffraction data analysis.
Figures
Similar articles
-
Blocking the shikimate pathway amplifies the impact of carvacrol on biofilm formation in Candida albicans.Microbiol Spectr. 2025 Mar 4;13(3):e0275424. doi: 10.1128/spectrum.02754-24. Epub 2025 Feb 7. Microbiol Spectr. 2025. PMID: 39918333 Free PMC article.
-
Candida albicans Aro1 affects cell wall integrity, biofilm formation and virulence.J Microbiol Immunol Infect. 2020 Feb;53(1):115-124. doi: 10.1016/j.jmii.2018.04.002. Epub 2018 May 17. J Microbiol Immunol Infect. 2020. PMID: 29807722
-
Glutathione biosynthesis in the yeast pathogens Candida glabrata and Candida albicans: essential in C. glabrata, and essential for virulence in C. albicans.Microbiology (Reading). 2011 Feb;157(Pt 2):484-495. doi: 10.1099/mic.0.045054-0. Epub 2010 Oct 21. Microbiology (Reading). 2011. PMID: 20966090
-
Candida and candidaemia. Susceptibility and epidemiology.Dan Med J. 2013 Nov;60(11):B4698. Dan Med J. 2013. PMID: 24192246 Review.
-
Caspofungin resistance in Candida albicans: genetic factors and synergistic compounds for combination therapies.Braz J Microbiol. 2022 Sep;53(3):1101-1113. doi: 10.1007/s42770-022-00739-9. Epub 2022 Mar 29. Braz J Microbiol. 2022. PMID: 35352319 Free PMC article. Review.
Cited by
-
Ent2 Governs Morphogenesis and Virulence in Part through Regulation of the Cdc42 Signaling Cascade in the Fungal Pathogen Candida albicans.mBio. 2023 Apr 25;14(2):e0343422. doi: 10.1128/mbio.03434-22. Epub 2023 Feb 21. mBio. 2023. PMID: 36809010 Free PMC article.
-
Blocking the shikimate pathway amplifies the impact of carvacrol on biofilm formation in Candida albicans.Microbiol Spectr. 2025 Mar 4;13(3):e0275424. doi: 10.1128/spectrum.02754-24. Epub 2025 Feb 7. Microbiol Spectr. 2025. PMID: 39918333 Free PMC article.
References
-
- Afonine PV, Grosse-Kunstleve RW, Echols N, Headd JJ, Moriarty NW, Mustyakimov M, Terwilliger TC, Urzhumtsev A, Zwart PH, Adams PD (2012) Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr D Biol Crystallogr 68: 352–367. 10.1107/S0907444912001308 - DOI - PMC - PubMed
-
- Blanco B, Prado V, Lence E, Otero JM, Garcia-Doval C, van Raaij MJ, Llamas-Saiz AL, Lamb H, Hawkins AR, González-Bello C (2013) Mycobacterium tuberculosis shikimate kinase inhibitors: Design and simulation studies of the catalytic turnover. J Am Chem Soc 135: 12366–12376. 10.1021/ja405853p - DOI - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources