

ON/OFF domains shape receptive field structure in mouse visual cortex
- PMID: 35513375
- PMCID: PMC9072422
- DOI: 10.1038/s41467-022-29999-7
ON/OFF domains shape receptive field structure in mouse visual cortex
Abstract
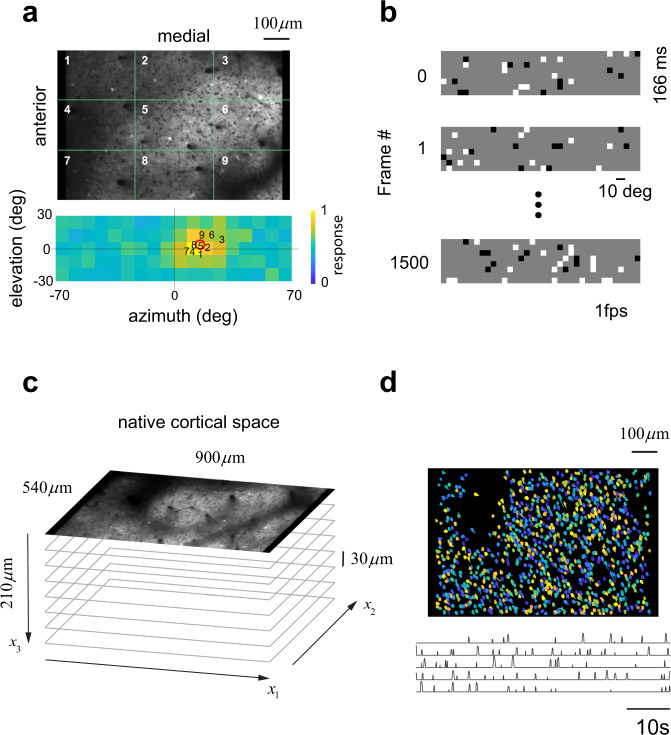
In higher mammals, thalamic afferents to primary visual cortex (area V1) segregate according to their responses to increases (ON) or decreases (OFF) in luminance. This organization induces columnar, ON/OFF domains postulated to provide a scaffold for the emergence of orientation tuning. To further test this idea, we asked whether ON/OFF domains exist in mouse V1. Here we show that mouse V1 is indeed parceled into ON/OFF domains. Interestingly, fluctuations in the relative density of ON/OFF neurons on the cortical surface mirror fluctuations in the relative density of ON/OFF receptive field centers on the visual field. Moreover, the local diversity of cortical receptive fields is explained by a model in which neurons linearly combine a small number of ON and OFF signals available in their cortical neighborhoods. These findings suggest that ON/OFF domains originate in fluctuations of the balance between ON/OFF responses across the visual field which, in turn, shapes the structure of cortical receptive fields.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
