

Rad51-mediated replication of damaged templates relies on monoSUMOylated DDK kinase
- PMID: 35513396
- PMCID: PMC9072374
- DOI: 10.1038/s41467-022-30215-9
Rad51-mediated replication of damaged templates relies on monoSUMOylated DDK kinase
Abstract
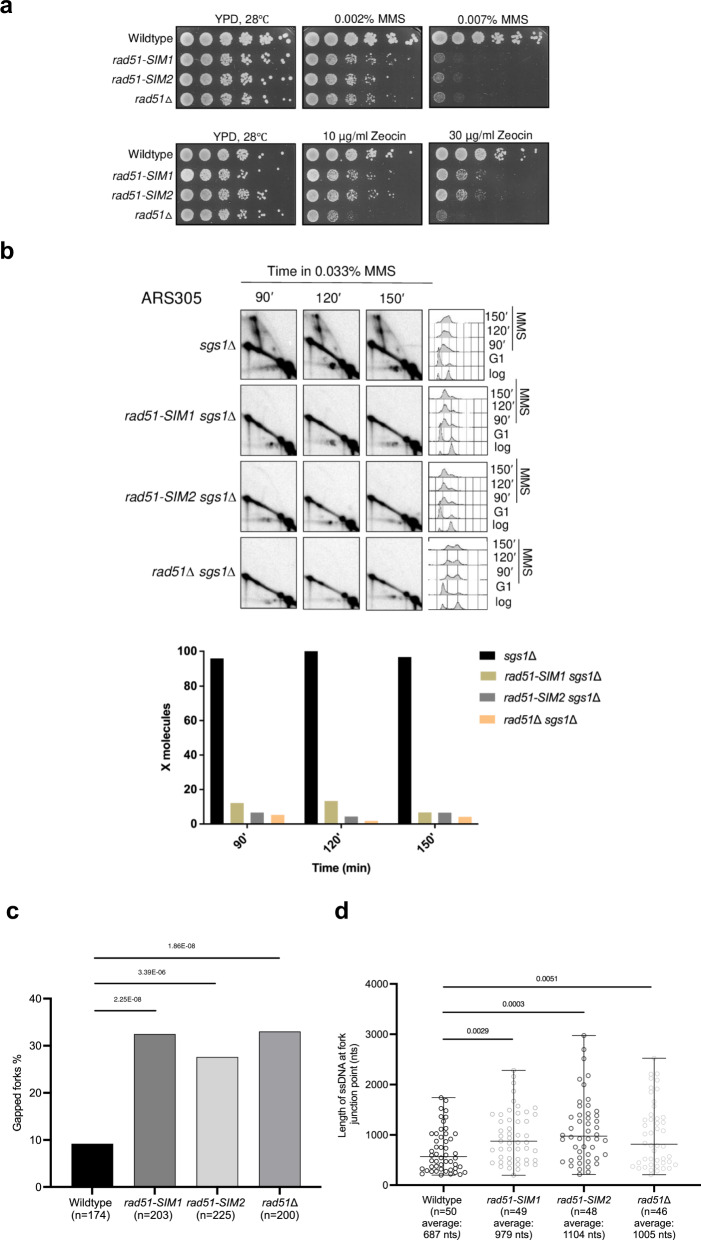
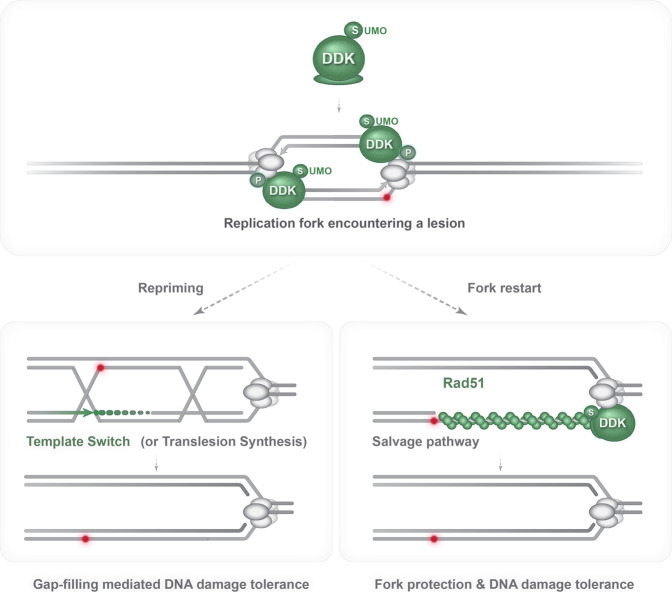
DNA damage tolerance (DDT), activated by replication stress during genome replication, is mediated by translesion synthesis and homologous recombination (HR). Here we uncover that DDK kinase, essential for replication initiation, is critical for replication-associated recombination-mediated DDT. DDK relies on its multi-monoSUMOylation to facilitate HR-mediated DDT and optimal retention of Rad51 recombinase at replication damage sites. Impairment of DDK kinase activity, reduced monoSUMOylation and mutations in the putative SUMO Interacting Motifs (SIMs) of Rad51 impair replication-associated recombination and cause fork uncoupling with accumulation of large single-stranded DNA regions at fork branching points. Notably, genetic activation of salvage recombination rescues the uncoupled fork phenotype but not the recombination-dependent gap-filling defect of DDK mutants, revealing that the salvage recombination pathway operates preferentially proximal to fork junctions at stalled replication forks. Overall, we uncover that monoSUMOylated DDK acts with Rad51 in an axis that prevents replication fork uncoupling and mediates recombination-dependent gap-filling.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
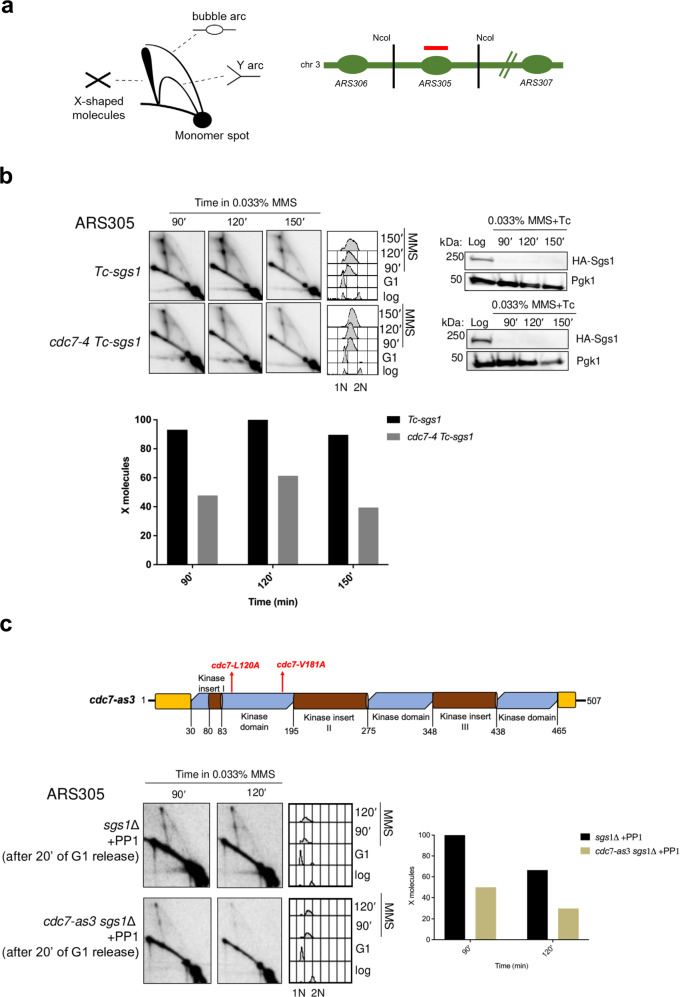
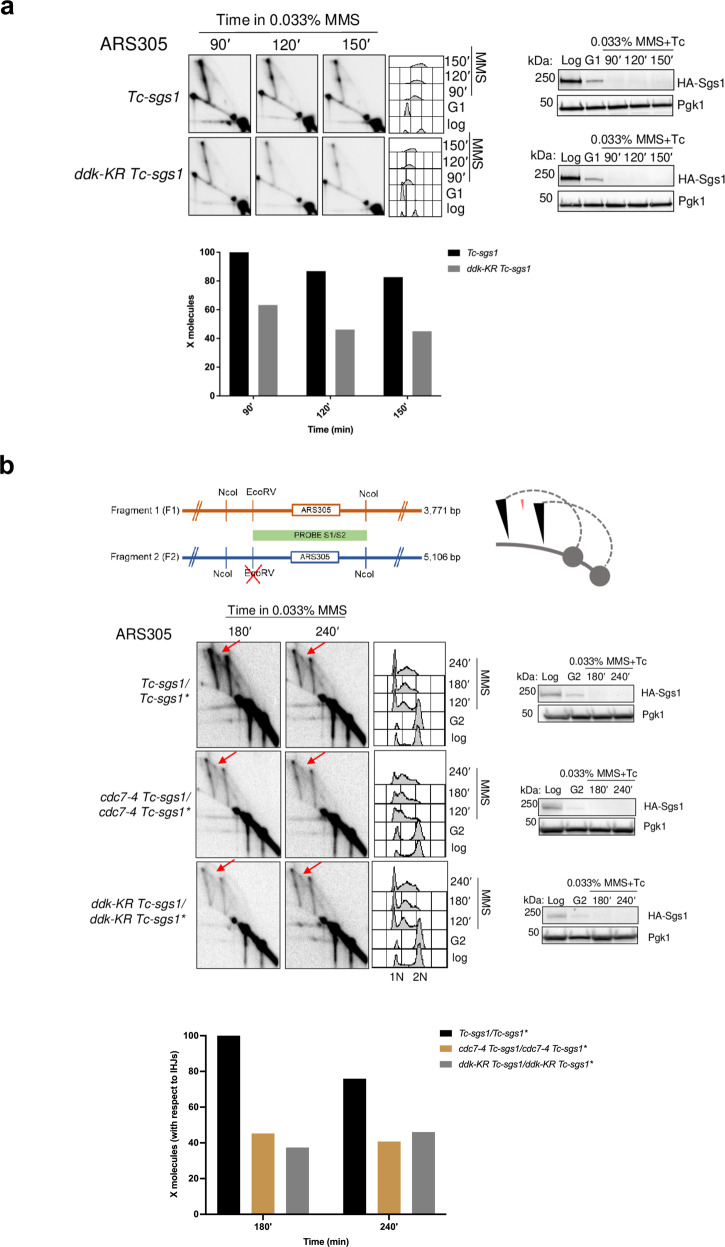
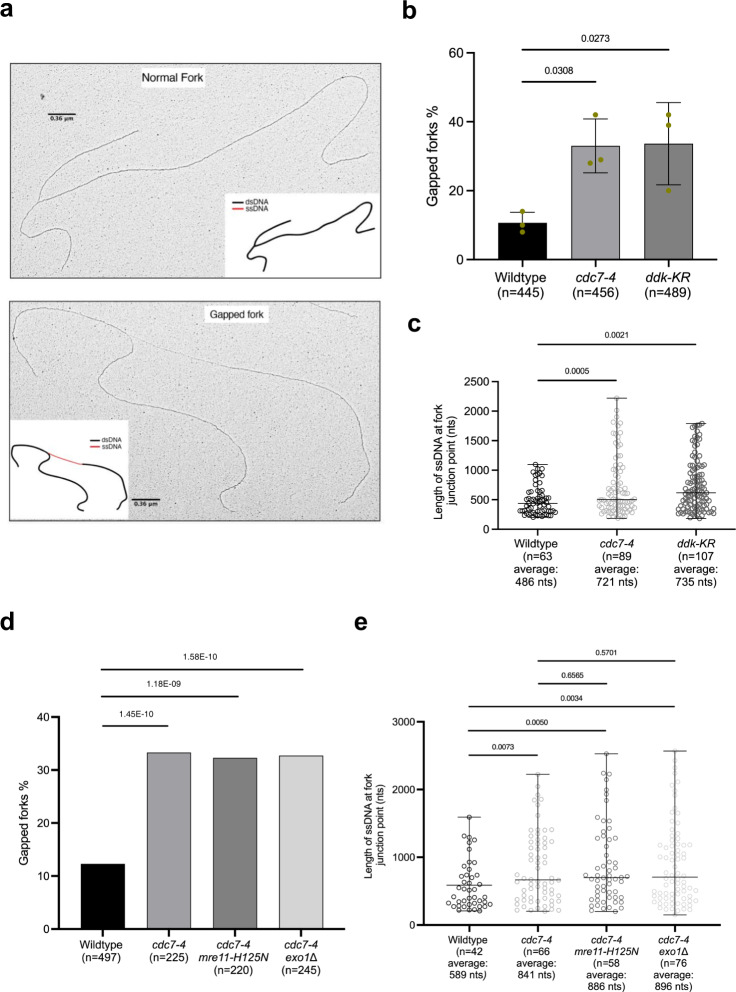
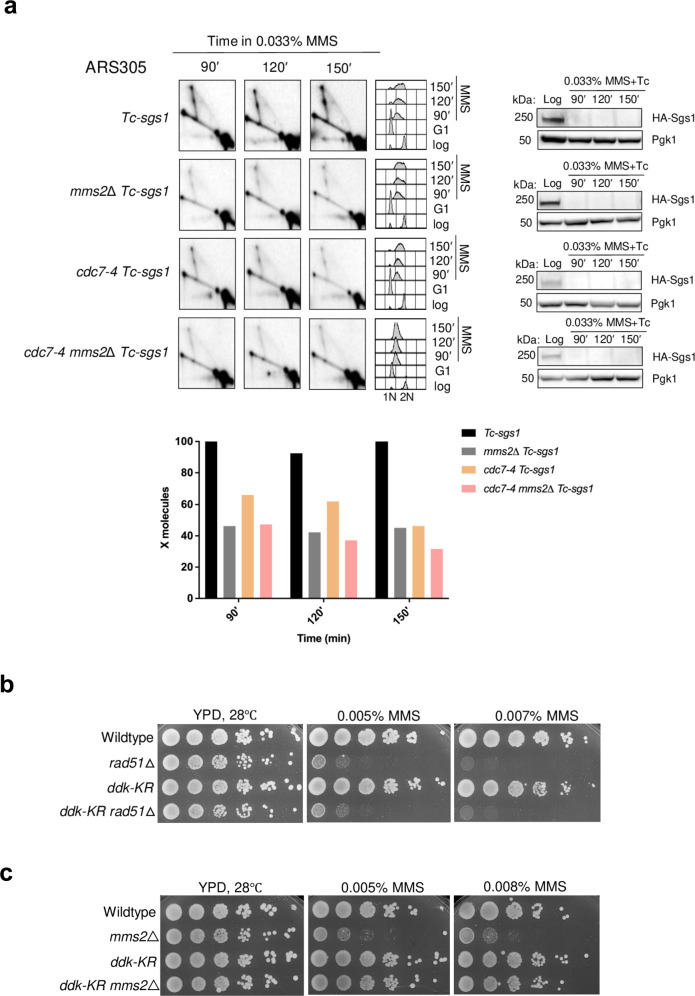

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
