

Spindle motility skews division site determination during asymmetric cell division in Physcomitrella
- PMID: 35513464
- PMCID: PMC9072379
- DOI: 10.1038/s41467-022-30239-1
Spindle motility skews division site determination during asymmetric cell division in Physcomitrella
Abstract
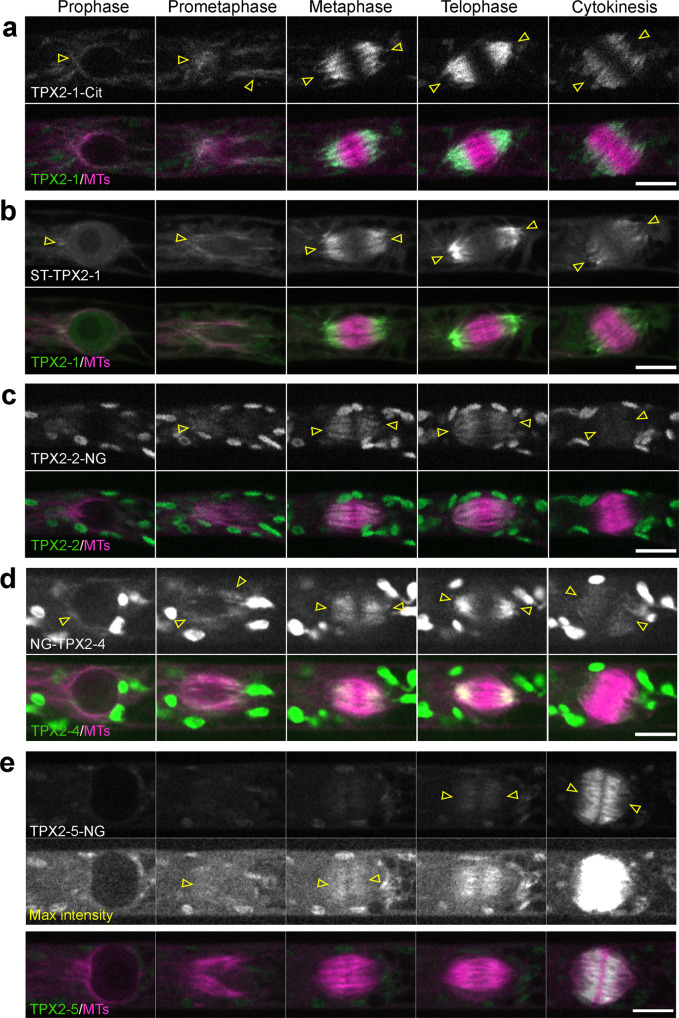
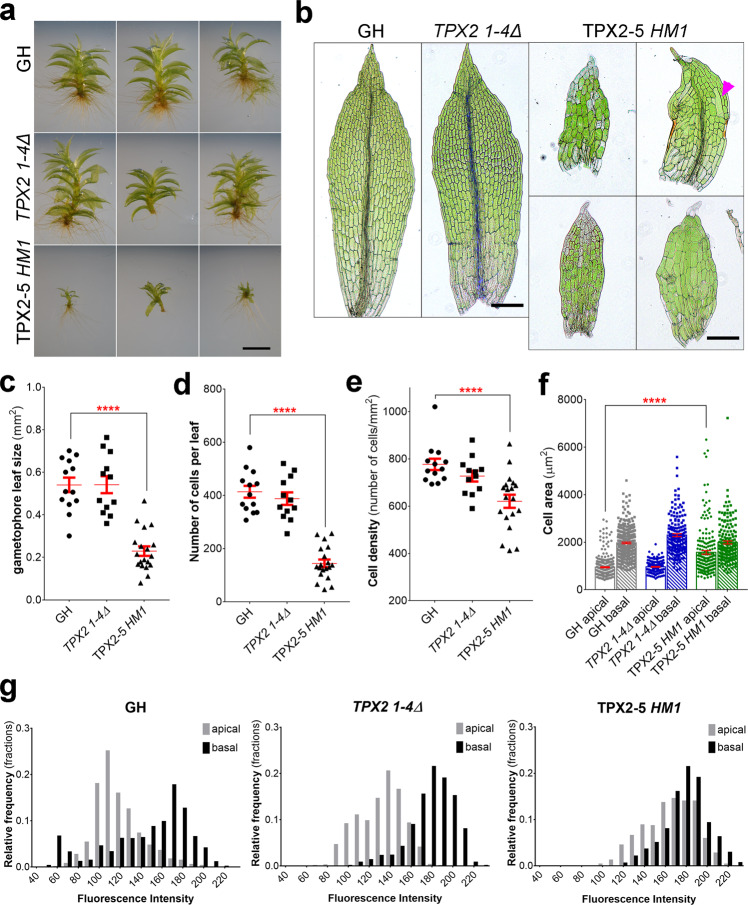
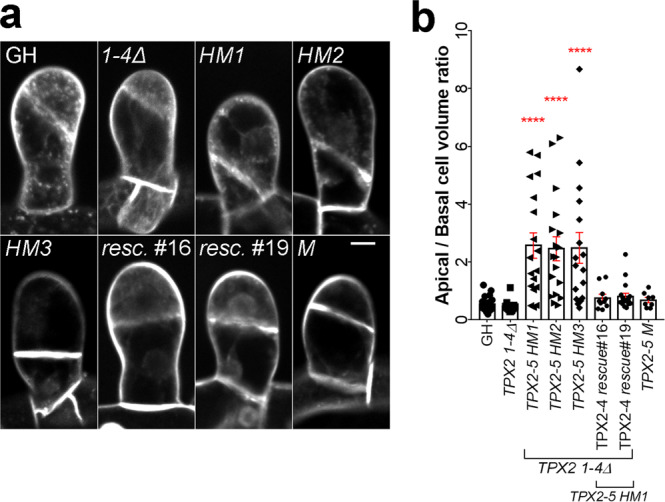
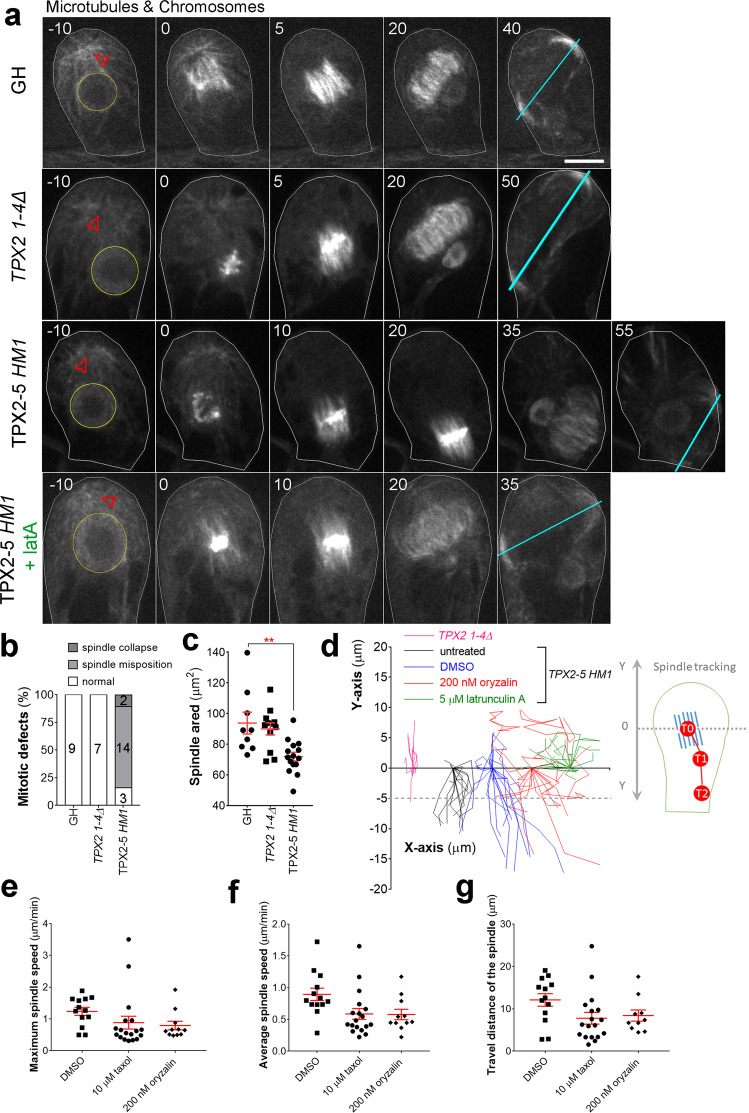
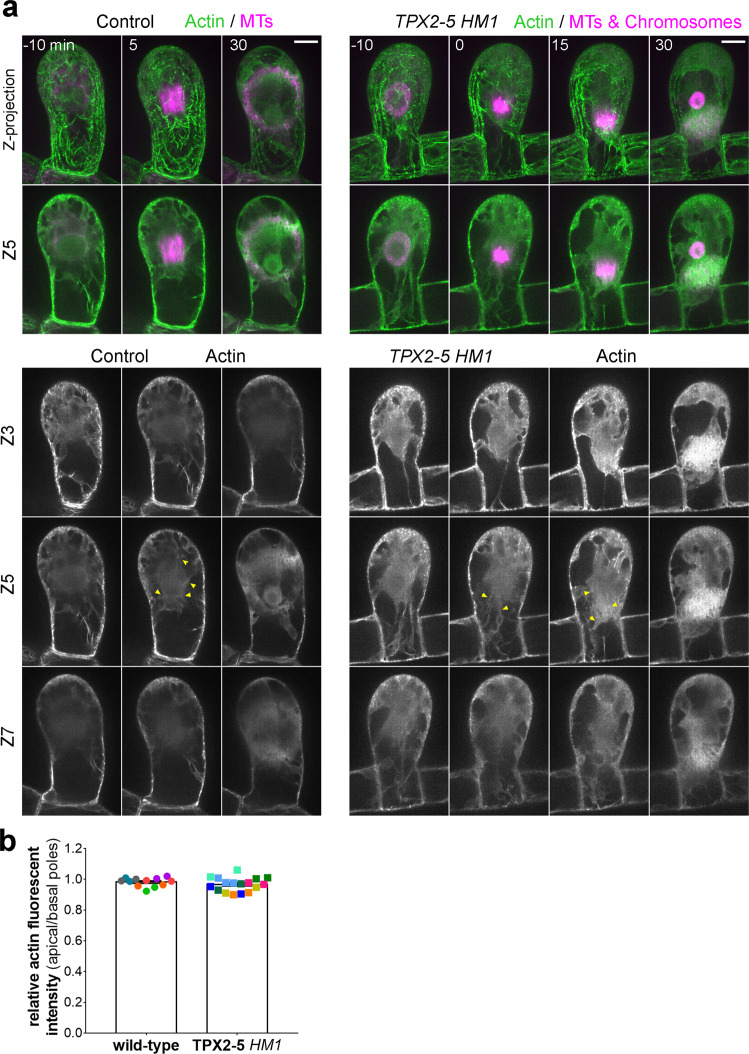
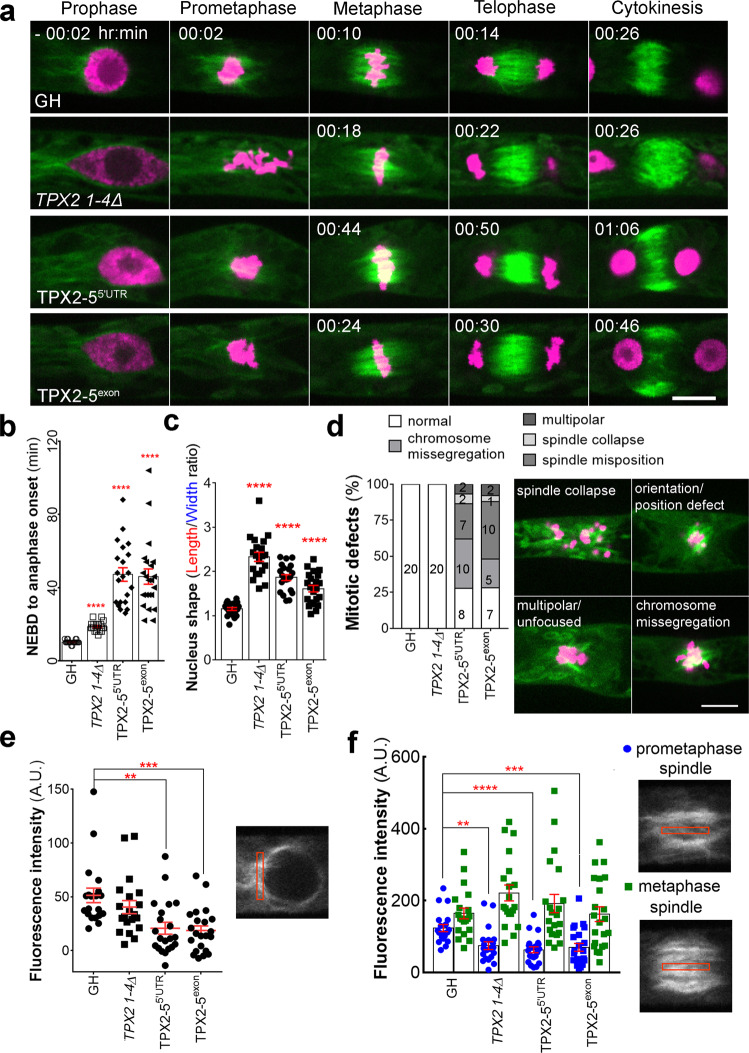
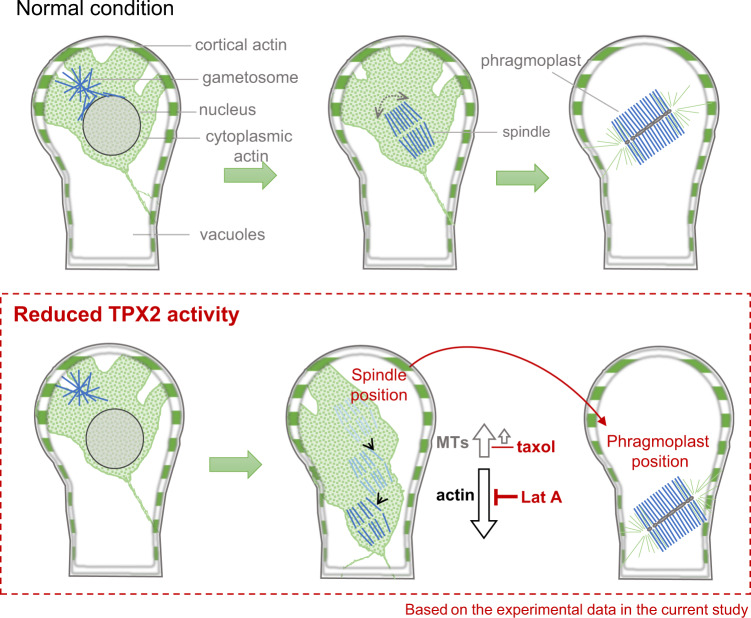
Asymmetric cell division (ACD) underlies the development of multicellular organisms. In animal ACD, the cell division site is determined by active spindle-positioning mechanisms. In contrast, it is considered that the division site in plants is determined prior to mitosis by the microtubule-actin belt known as the preprophase band (PPB) and that the localization of the mitotic spindle is typically static and does not govern the division plane. However, in some plant species, ACD occurs in the absence of PPB. Here, we isolate a hypomorphic mutant of the conserved microtubule-associated protein TPX2 in the moss Physcomitrium patens (Physcomitrella) and observe spindle motility during PPB-independent cell division. This defect compromises the position of the division site and produces inverted daughter cell sizes in the first ACD of gametophore (leafy shoot) development. The phenotype is rescued by restoring endogenous TPX2 function and, unexpectedly, by depolymerizing actin filaments. Thus, we identify an active spindle-positioning mechanism that, reminiscent of acentrosomal ACD in animals, involves microtubules and actin filaments, and sets the division site in plants.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Cytoplasmic MTOCs control spindle orientation for asymmetric cell division in plants.Proc Natl Acad Sci U S A. 2017 Oct 17;114(42):E8847-E8854. doi: 10.1073/pnas.1713925114. Epub 2017 Oct 2. Proc Natl Acad Sci U S A. 2017. PMID: 28973935 Free PMC article.
-
Myosin XI localizes at the mitotic spindle and along the cell plate during plant cell division in Physcomitrella patens.Biochem Biophys Res Commun. 2018 Nov 25;506(2):409-421. doi: 10.1016/j.bbrc.2018.01.082. Epub 2018 Jan 12. Biochem Biophys Res Commun. 2018. PMID: 29339158
-
The function of TONNEAU1 in moss reveals ancient mechanisms of division plane specification and cell elongation in land plants.Development. 2010 Aug;137(16):2733-42. doi: 10.1242/dev.043810. Development. 2010. PMID: 20663817
-
The role of the cytoskeleton and associated proteins in determination of the plant cell division plane.Plant J. 2013 Jul;75(2):258-69. doi: 10.1111/tpj.12177. Epub 2013 Apr 19. Plant J. 2013. PMID: 23496276 Review.
-
Multifaceted regulation of asymmetric cell division by the actin cytoskeleton.Curr Opin Cell Biol. 2025 Jun;94:102491. doi: 10.1016/j.ceb.2025.102491. Epub 2025 Mar 7. Curr Opin Cell Biol. 2025. PMID: 40056852 Review.
Cited by
-
CURLY LEAF is required for the auxin-dependent regulation of 3-dimensional growth specification in Physcomitrium patens.MicroPubl Biol. 2023 Apr 17;2023:10.17912/micropub.biology.000797. doi: 10.17912/micropub.biology.000797. eCollection 2023. MicroPubl Biol. 2023. PMID: 37143449 Free PMC article.
-
HDAC6 deacetylates ENKD1 to regulate mitotic spindle behavior and corneal epithelial homeostasis.EMBO Rep. 2025 May;26(10):2597-2621. doi: 10.1038/s44319-025-00438-0. Epub 2025 Mar 28. EMBO Rep. 2025. PMID: 40155750 Free PMC article.
-
Cell size: a key determinant of meristematic potential in plant protoplasts.aBIOTECH. 2021 Jan 1;2(1):96-104. doi: 10.1007/s42994-020-00033-y. eCollection 2021 Mar. aBIOTECH. 2021. PMID: 36304480 Free PMC article. Review.
-
The TOPLESS corepressor regulates developmental switches in the bryophyte Physcomitrium patens that were critical for plant terrestrialisation.Plant J. 2023 Sep;115(5):1331-1344. doi: 10.1111/tpj.16322. Epub 2023 Jun 8. Plant J. 2023. PMID: 37243383 Free PMC article.
-
Plant stem cell research is uncovering the secrets of longevity and persistent growth.Plant J. 2021 Apr;106(2):326-335. doi: 10.1111/tpj.15184. Epub 2021 Mar 25. Plant J. 2021. PMID: 33533118 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
