

Side-by-side comparison of Notch- and C83 binding to γ-secretase in a complete membrane model at physiological temperature
- PMID: 35520661
- PMCID: PMC9056423
- DOI: 10.1039/d0ra04683c
Side-by-side comparison of Notch- and C83 binding to γ-secretase in a complete membrane model at physiological temperature
Abstract
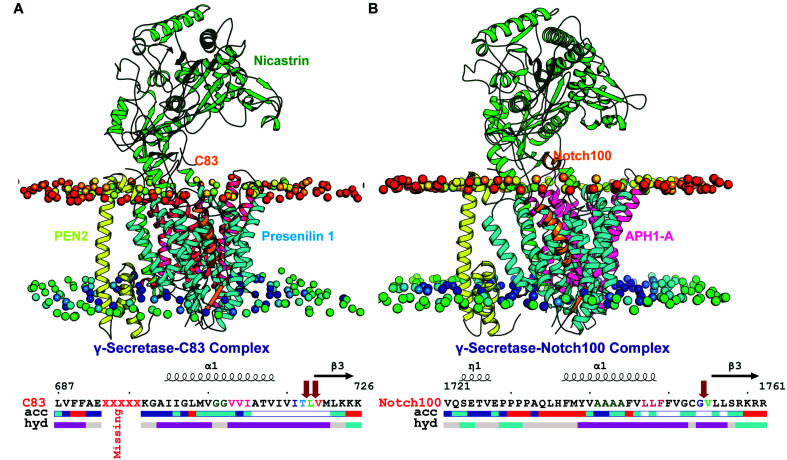
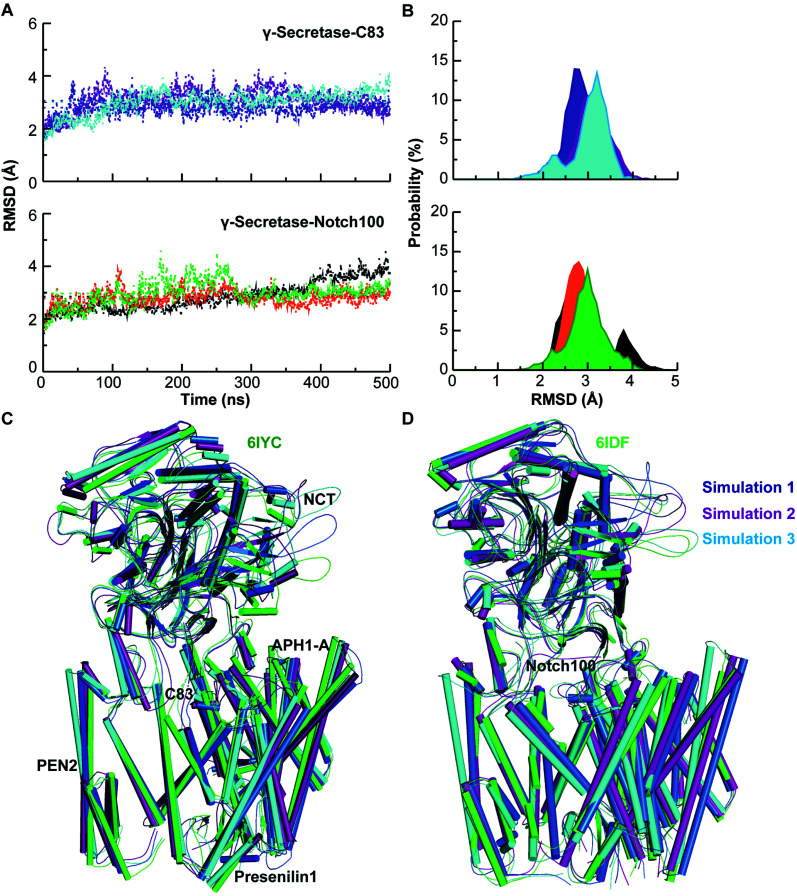
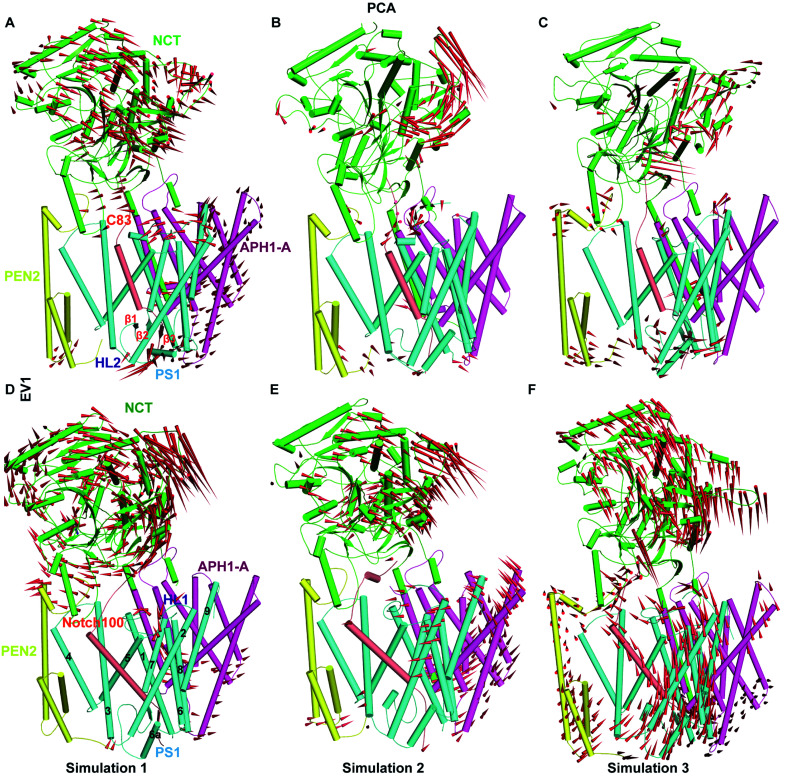
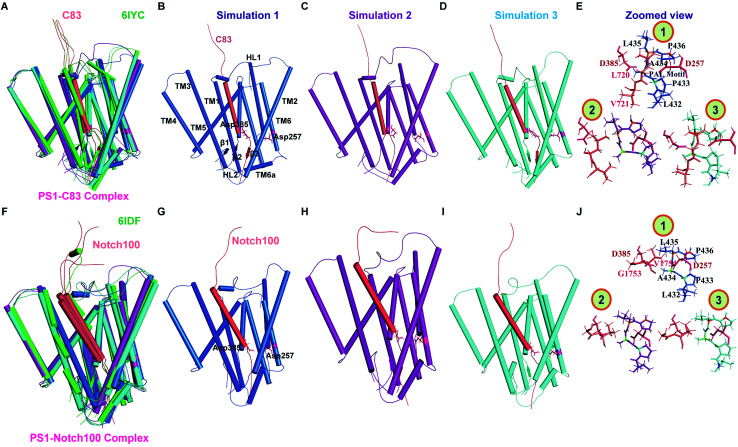
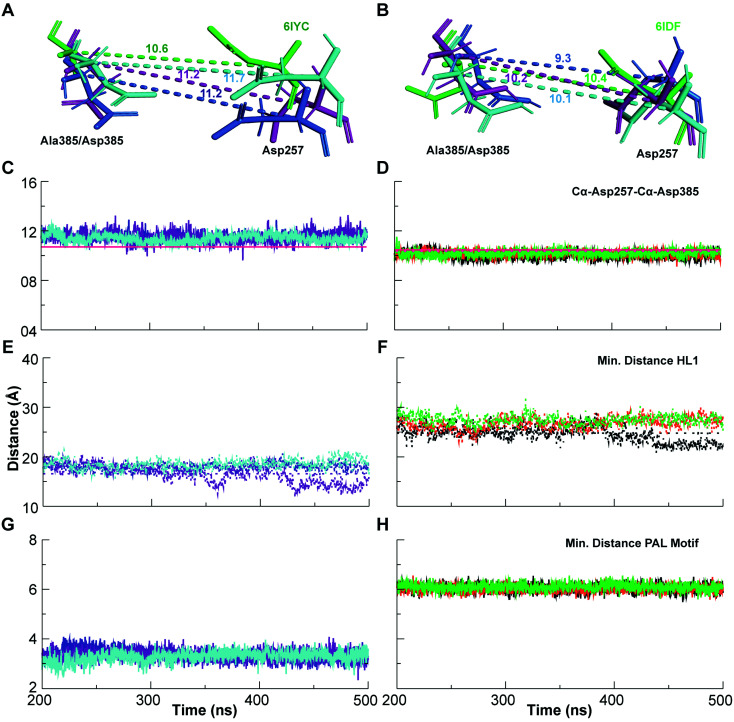
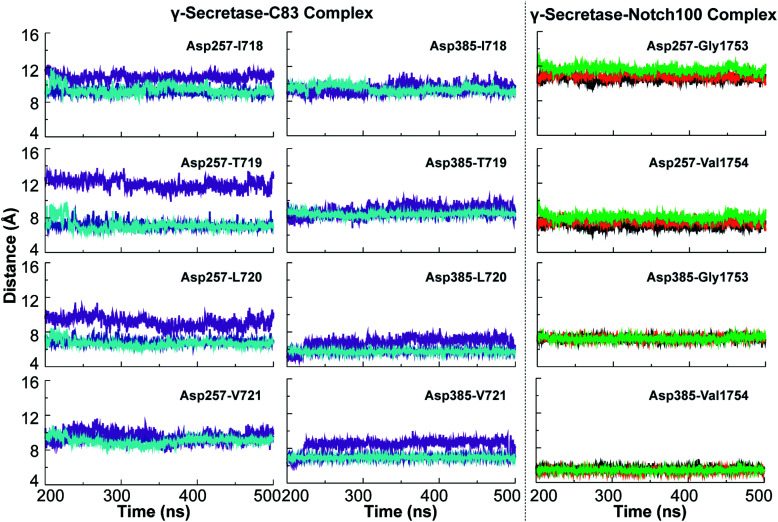
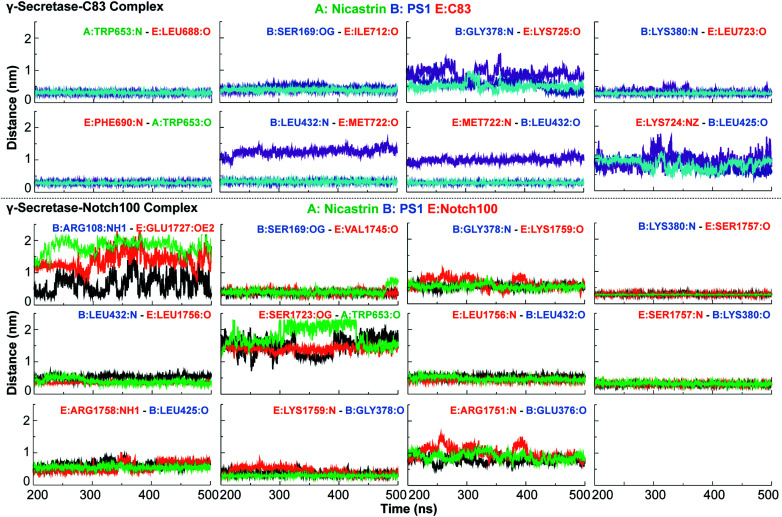
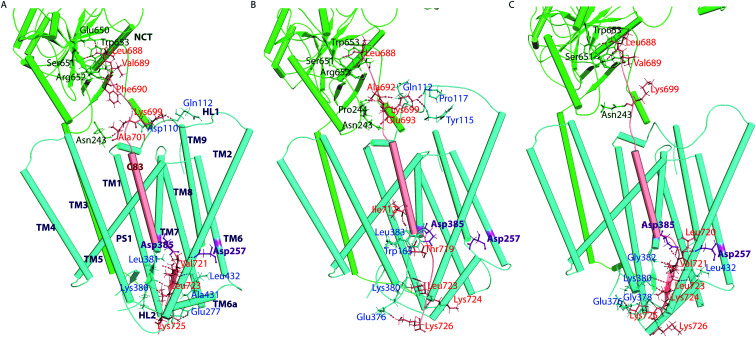
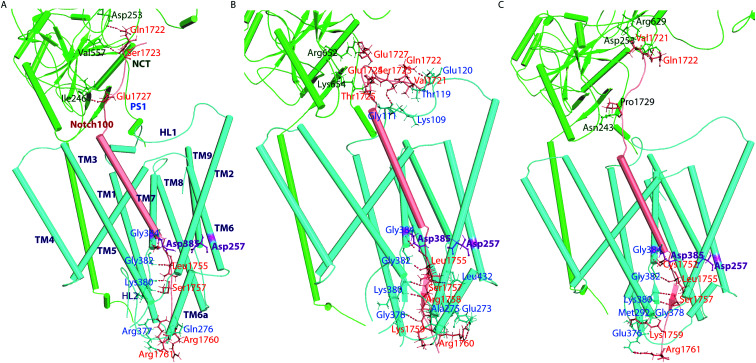
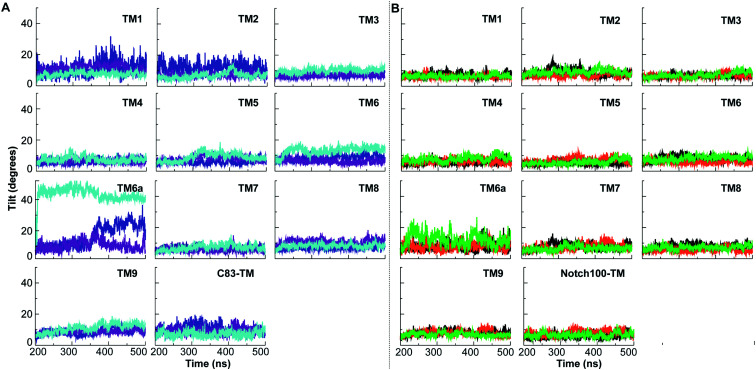
γ-Secretase cleaves the C99 fragment of the amyloid precursor protein, leading to formation of aggregated β-amyloid peptide central to Alzheimer's disease, and Notch, essential for cell regulation. Recent cryogenic electron microscopy (cryo-EM) structures indicate major changes upon substrate binding, a β-sheet recognition motif, and a possible helix unwinding to expose peptide bonds towards nucleophilic attack. Here we report side-by-side comparison of the 303 K dynamics of the two proteins in realistic membranes using molecular dynamics simulations. Our ensembles agree with the cryo-EM data (full-protein Cα-RMSD = 1.62-2.19 Å) but reveal distinct presenilin helix conformation states and thermal β-strand to coil transitions of C83 and Notch100. We identify distinct 303 K hydrogen bond dynamics and water accessibility of the catalytic sites. The RKRR motif (1758-1761) contributes significantly to Notch binding and serves as a "membrane anchor" that prevents Notch displacement. Water that transiently hydrogen bonds to G1753 and V1754 probably represents the catalytic nucleophile. At 303 K, Notch and C83 binding induce different conformation states, with Notch mostly present in a closed state with shorter Asp-Asp distance. This may explain the different outcome of Notch and C99 cleavage, as the latter is more imprecise with many products. Our identified conformation states may aid efforts to develop conformation-selective drugs that target C99 and Notch cleavage differently, e.g. Notch-sparing γ-secretase modulators.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
All authors hereby declare that they have no competing interests, neither financial nor non-financial, related to this work.
Figures
Similar articles
-
Molecular dynamics of C99-bound γ-secretase reveal two binding modes with distinct compactness, stability, and active-site retention: implications for Aβ production.Biochem J. 2019 Apr 15;476(7):1173-1189. doi: 10.1042/BCJ20190023. Biochem J. 2019. PMID: 30910800
-
Membrane dynamics of γ-secretase with the anterior pharynx-defective 1B subunit.J Cell Biochem. 2021 Jan;122(1):69-85. doi: 10.1002/jcb.29832. Epub 2020 Aug 23. J Cell Biochem. 2021. PMID: 32830360
-
A computer-simulated mechanism of familial Alzheimer's disease: Mutations enhance thermal dynamics and favor looser substrate-binding to γ-secretase.J Struct Biol. 2020 Dec 1;212(3):107648. doi: 10.1016/j.jsb.2020.107648. Epub 2020 Oct 21. J Struct Biol. 2020. PMID: 33099014
-
Substrate-Enzyme Interactions in Intramembrane Proteolysis: γ-Secretase as the Prototype.Front Mol Neurosci. 2020 May 19;13:65. doi: 10.3389/fnmol.2020.00065. eCollection 2020. Front Mol Neurosci. 2020. PMID: 32508589 Free PMC article. Review.
-
Insight γ-Secretase: Structure, Function, and Role in Alzheimer's Disease.Curr Drug Targets. 2021;22(12):1376-1403. doi: 10.2174/1389450121999201230203709. Curr Drug Targets. 2021. PMID: 33390127 Review.
Cited by
-
MDM-TASK-web: MD-TASK and MODE-TASK web server for analyzing protein dynamics.Comput Struct Biotechnol J. 2021 Sep 2;19:5059-5071. doi: 10.1016/j.csbj.2021.08.043. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 34589183 Free PMC article.
-
Effects of presenilin-1 familial Alzheimer's disease mutations on γ-secretase activation for cleavage of amyloid precursor protein.Commun Biol. 2023 Feb 14;6(1):174. doi: 10.1038/s42003-023-04539-1. Commun Biol. 2023. PMID: 36788318 Free PMC article.
-
Formation of extramembrane β-strands controls dimerization of transmembrane helices in amyloid precursor protein C99.Proc Natl Acad Sci U S A. 2022 Dec 27;119(52):e2212207119. doi: 10.1073/pnas.2212207119. Epub 2022 Dec 20. Proc Natl Acad Sci U S A. 2022. PMID: 36538482 Free PMC article.
-
Sequence-structure functional implications and molecular simulation of high deleterious nonsynonymous substitutions in IDH1 revealed the mechanism of drug resistance in glioma.Front Pharmacol. 2022 Sep 16;13:927570. doi: 10.3389/fphar.2022.927570. eCollection 2022. Front Pharmacol. 2022. PMID: 36188571 Free PMC article.
-
Mechanism of Tripeptide Trimming of Amyloid β-Peptide 49 by γ-Secretase.J Am Chem Soc. 2022 Apr 13;144(14):6215-6226. doi: 10.1021/jacs.1c10533. Epub 2022 Apr 4. J Am Chem Soc. 2022. PMID: 35377629 Free PMC article.
References
-
- Nichols E. Szoeke C. E. I. Vollset S. E. Abbasi N. Abd-Allah F. Abdela J. Aichour M. T. E. Akinyemi R. O. Alahdab F. Asgedom S. W. Awasthi A. Barker-Collo S. L. Baune B. T. Béjot Y. Belachew A. B. Bennett D. A. Biadgo B. Bijani A. Bin Sayeed M. S. Brayne C. Carpenter D. O. Carvalho F. Catalá-López F. Cerin E. Choi J. Y. J. Dang A. K. Degefa M. G. Djalalinia S. Dubey M. Duken E. E. Edvardsson D. Endres M. Eskandarieh S. Faro A. Farzadfar F. Fereshtehnejad S. M. Fernandes E. Filip I. Fischer F. Gebre A. K. Geremew D. Ghasemi-Kasman M. Gnedovskaya E. V. Gupta R. Hachinski V. Hagos T. B. Hamidi S. Hankey G. J. Haro J. M. Hay S. I. Irvani S. S. N. Jha R. P. Jonas J. B. Kalani R. Karch A. Kasaeian A. Khader Y. S. Khalil I. A. Khan E. A. Khanna T. Khoja T. A. M. Khubchandani J. Kisa A. Kissimova-Skarbek K. Kivimäki M. Koyanagi A. Krohn K. J. Logroscino G. Lorkowski S. Majdan M. Malekzadeh R. März W. Massano J. Mengistu G. Meretoja A. Mohammadi M. Mohammadi-Khanaposhtani M. Mokdad A. H. Mondello S. Moradi G. Nagel G. Naghavi M. Naik G. Nguyen L. H. Nguyen T. H. Nirayo Y. L. Nixon M. R. Ofori-Asenso R. Ogbo F. A. Olagunju A. T. Owolabi M. O. Panda-Jonas S. Passos V. M. d. A. Pereira D. M. Pinilla-Monsalve G. D. Piradov M. A. Pond C. D. Poustchi H. Qorbani M. Radfar A. Reiner R. C. Robinson S. R. Roshandel G. Rostami A. Russ T. C. Sachdev P. S. Safari H. Safiri S. Sahathevan R. Salimi Y. Satpathy M. Sawhney M. Saylan M. Sepanlou S. G. Shafieesabet A. Shaikh M. A. Sahraian M. A. Shigematsu M. Shiri R. Shiue I. Silva J. P. Smith M. Sobhani S. Stein D. J. Tabarés-Seisdedos R. Tovani-Palone M. R. Tran B. X. Tran T. T. Tsegay A. T. Ullah I. Venketasubramanian N. Vlassov V. Wang Y. P. Weiss J. Westerman R. Wijeratne T. Wyper G. M. A. Yano Y. Yimer E. M. Yonemoto N. Yousefifard M. Zaidi Z. Zare Z. Vos T. Feigin V. L. Murray C. J. L. Lancet Neurol. 2019;18:88–106. doi: 10.1016/S1474-4422(18)30403-4. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources