

The HD-Zip transcription factor SlHB15A regulates abscission by modulating jasmonoyl-isoleucine biosynthesis
- PMID: 35522030
- PMCID: PMC9342995
- DOI: 10.1093/plphys/kiac212
The HD-Zip transcription factor SlHB15A regulates abscission by modulating jasmonoyl-isoleucine biosynthesis
Abstract
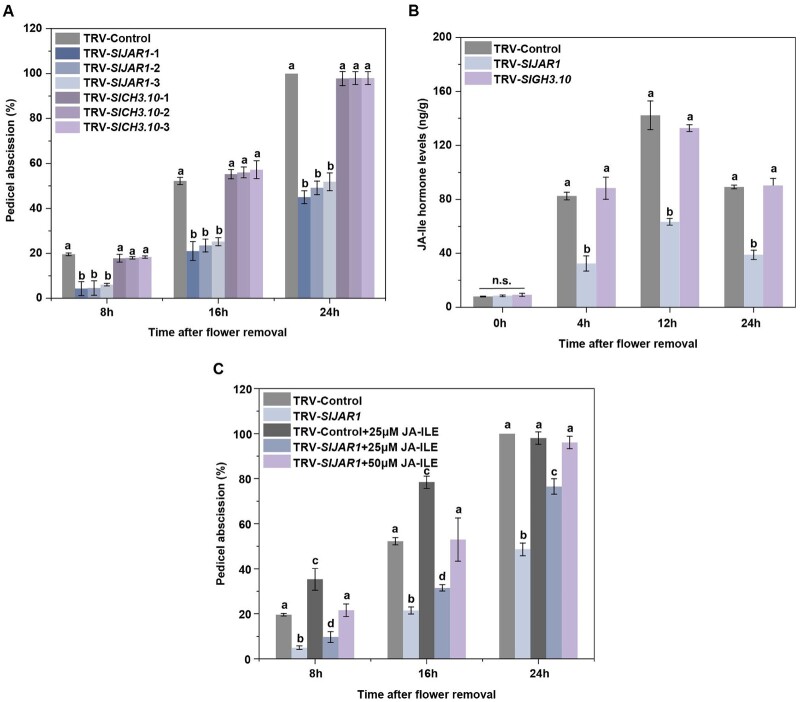
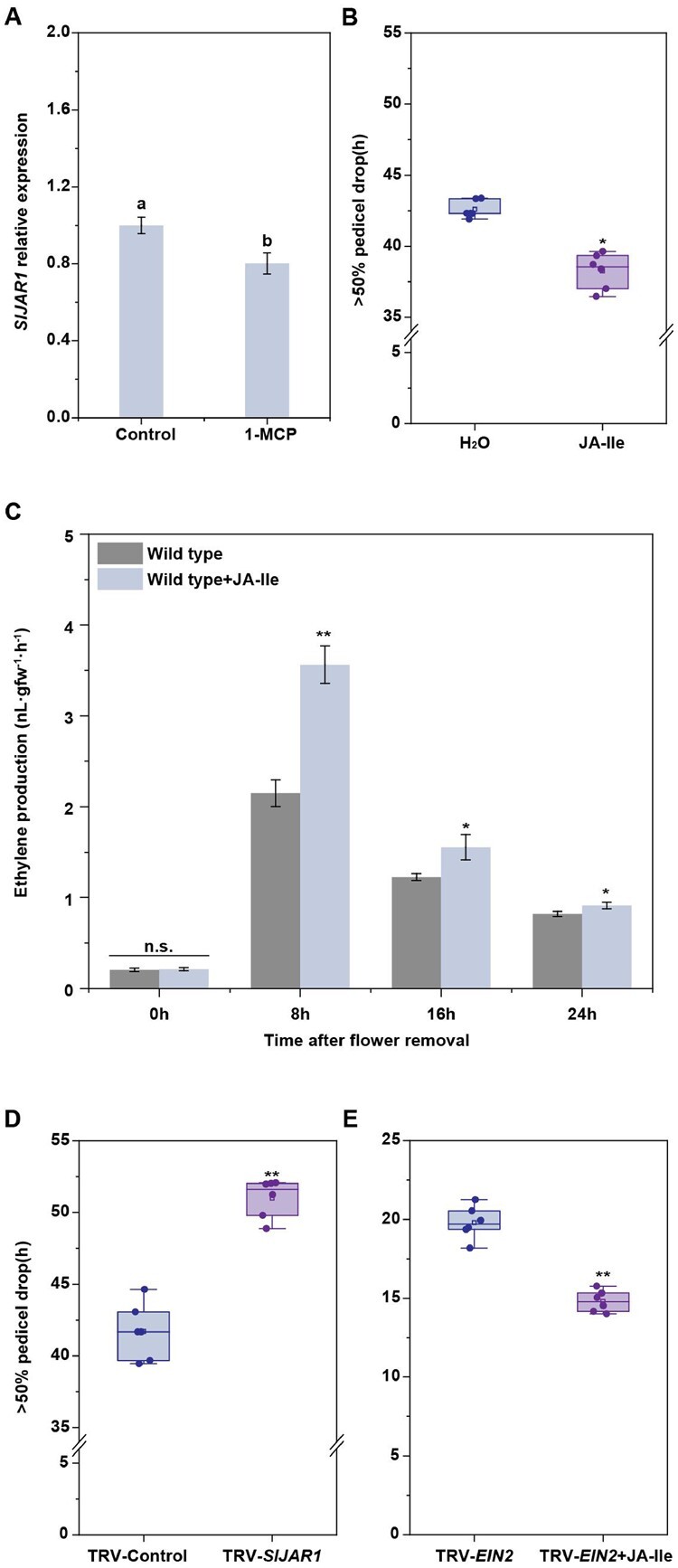
Plant organ abscission, a process that is important for development and reproductive success, is inhibited by the phytohormone auxin and promoted by another phytohormone, jasmonic acid (JA). However, the molecular mechanisms underlying the antagonistic effects of auxin and JA in organ abscission are unknown. We identified a tomato (Solanum lycopersicum) class III homeodomain-leucine zipper transcription factor, HOMEOBOX15A (SlHB15A), which was highly expressed in the flower pedicel abscission zone and induced by auxin. Knocking out SlHB15A using clustered regularly interspaced short palindromic repeats-associated protein 9 technology significantly accelerated abscission. In contrast, overexpression of microRNA166-resistant SlHB15A (mSlHB15A) delayed abscission. RNA sequencing and reverse transcription-quantitative PCR analyses showed that knocking out SlHB15A altered the expression of genes related to JA biosynthesis and signaling. Furthermore, functional analysis indicated that SlHB15A regulates abscission by depressing JA-isoleucine (JA-Ile) levels through inhabiting the expression of JASMONATE-RESISTANT1 (SlJAR1), a gene involved in JA-Ile biosynthesis, which could induce abscission-dependent and abscission-independent ethylene signaling. SlHB15A bound directly to the SlJAR1 promoter to silence SlJAR1, thus delaying abscission. We also found that flower removal enhanced JA-Ile content and that application of JA-Ile severely impaired the inhibitory effects of auxin on abscission. These results indicated that SlHB15A mediates the antagonistic effect of auxin and JA-Ile during tomato pedicel abscission, while auxin inhibits abscission through the SlHB15A-SlJAR1 module.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures







Similar articles
-
Tomato MYB21 Acts in Ovules to Mediate Jasmonate-Regulated Fertility.Plant Cell. 2019 May;31(5):1043-1062. doi: 10.1105/tpc.18.00978. Epub 2019 Mar 20. Plant Cell. 2019. PMID: 30894458 Free PMC article.
-
Ripening-induced defence signalling in Botrytis cinerea-infected tomato fruits involves activation of ERF.F4 by a MYC2-NOR/RIN protein complex.Plant Biotechnol J. 2025 Sep;23(9):4126-4139. doi: 10.1111/pbi.70221. Epub 2025 Jun 26. Plant Biotechnol J. 2025. PMID: 40568857 Free PMC article.
-
A small molecule antagonizes jasmonic acid perception and auxin responses in vascular and nonvascular plants.Plant Physiol. 2021 Nov 3;187(3):1399-1413. doi: 10.1093/plphys/kiab369. Plant Physiol. 2021. PMID: 34618088 Free PMC article.
-
Perception, signaling and cross-talk of jasmonates and the seminal contributions of the Daoxin Xie's lab and the Chuanyou Li's lab.Plant Cell Rep. 2014 May;33(5):707-18. doi: 10.1007/s00299-014-1608-5. Epub 2014 Apr 2. Plant Cell Rep. 2014. PMID: 24691578 Review.
-
Jasmonates in flower and seed development.Biochimie. 2013 Jan;95(1):79-85. doi: 10.1016/j.biochi.2012.06.005. Epub 2012 Jun 13. Biochimie. 2013. PMID: 22705387 Review.
Cited by
-
Abscission zone metabolism impacts pre- and post-harvest fruit quality: a very attaching story.Front Plant Sci. 2025 Jan 27;15:1524893. doi: 10.3389/fpls.2024.1524893. eCollection 2024. Front Plant Sci. 2025. PMID: 39980759 Free PMC article. Review.
-
Jarin-1, an inhibitor of JA-Ile biosynthesis in Arabidopsis thaliana, acts differently in other plant species.Plant Signal Behav. 2023 Dec 31;18(1):2273515. doi: 10.1080/15592324.2023.2273515. Epub 2023 Oct 30. Plant Signal Behav. 2023. PMID: 37902262 Free PMC article.
-
The Quantification of Jasmonates in Lipid-Rich Tissues by UHPLC-ESI-QQQ-MS/MS in Multiple Reaction Monitoring (MRM).Methods Mol Biol. 2025;2916:25-38. doi: 10.1007/978-1-0716-4470-6_3. Methods Mol Biol. 2025. PMID: 40366583
-
Genome-wide characterisation of HD-Zip transcription factors and functional analysis of PbHB24 during stone cell formation in Chinese white pear (Pyrus bretschneideri).BMC Plant Biol. 2024 May 23;24(1):444. doi: 10.1186/s12870-024-05138-w. BMC Plant Biol. 2024. PMID: 38778247 Free PMC article.
-
Abscission in plants: from mechanism to applications.Adv Biotechnol (Singap). 2024 Aug 9;2(3):27. doi: 10.1007/s44307-024-00033-9. Adv Biotechnol (Singap). 2024. PMID: 39883313 Free PMC article. Review.
References
-
- Baima S, Forte V, Possenti M, Peñalosa A, Leoni G, Salvi S, Felici B, Ruberti I, Morelli G (2014) Negative feedback regulation of auxin signaling by ATHB8/ACL5–BUD2 transcription module. Mol Plant 7:1006–1025 - PubMed
-
- Bozorov TA, Dinh ST, Baldwin IT (2017) JA but not JA‐Ile is the cell‐nonautonomous signal activating JA mediated systemic defenses to herbivory in Nicotiana attenuata. J Integr Plant Biol 59:552–571 - PubMed
