

Antigen presentation safeguards the integrity of the hematopoietic stem cell pool
- PMID: 35523139
- PMCID: PMC9202612
- DOI: 10.1016/j.stem.2022.04.007
Antigen presentation safeguards the integrity of the hematopoietic stem cell pool
Abstract
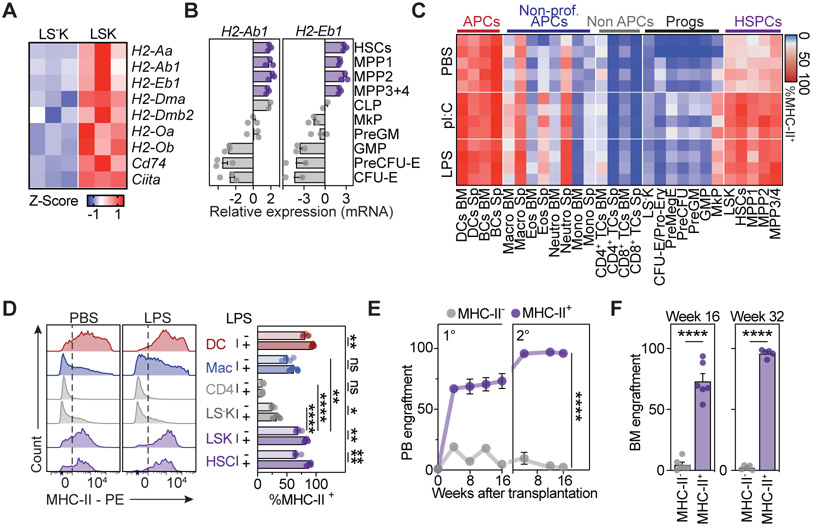
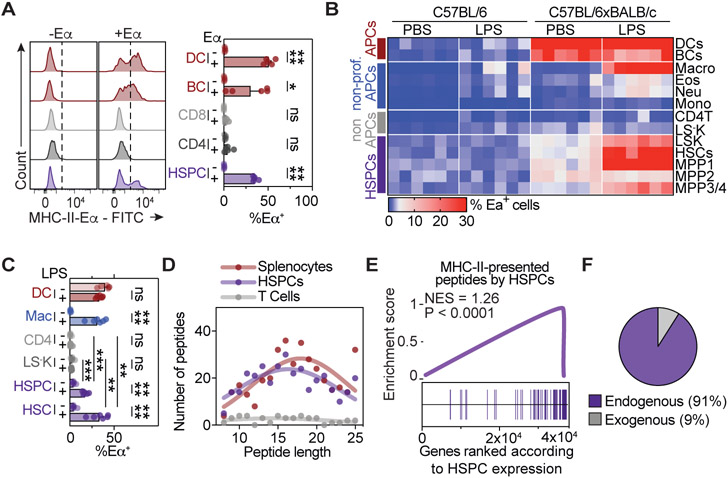
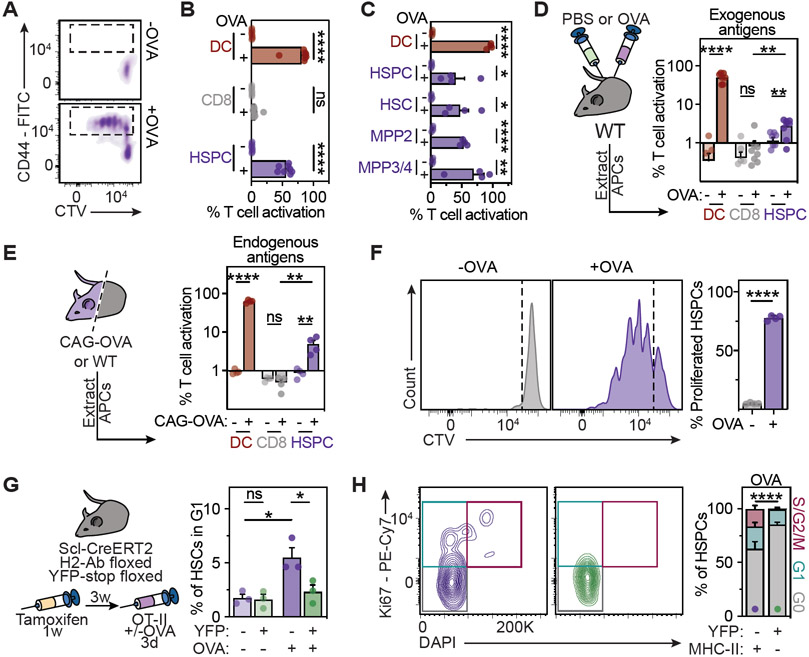
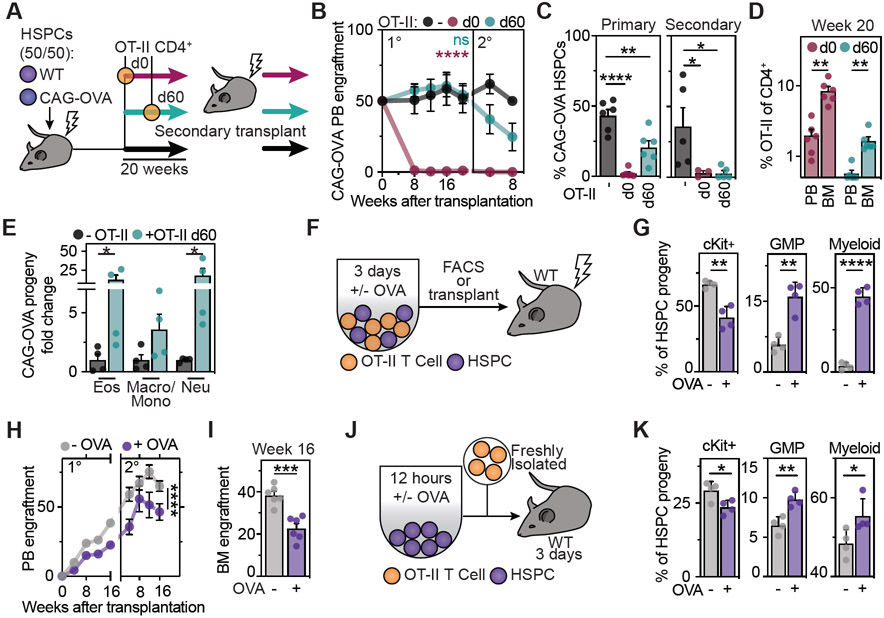
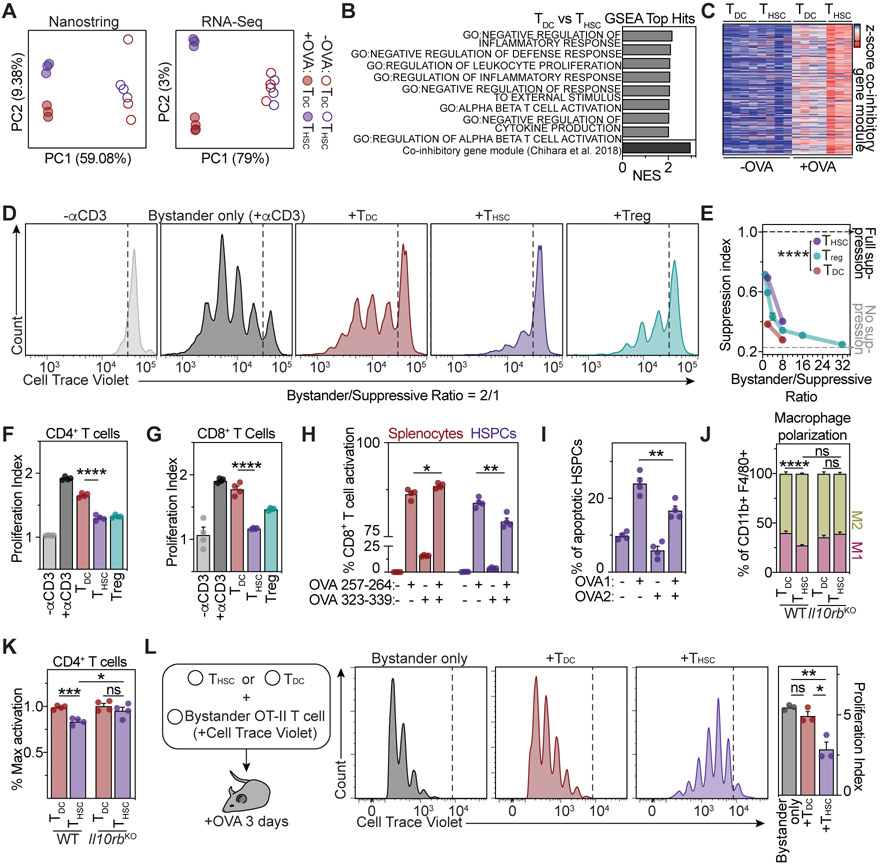
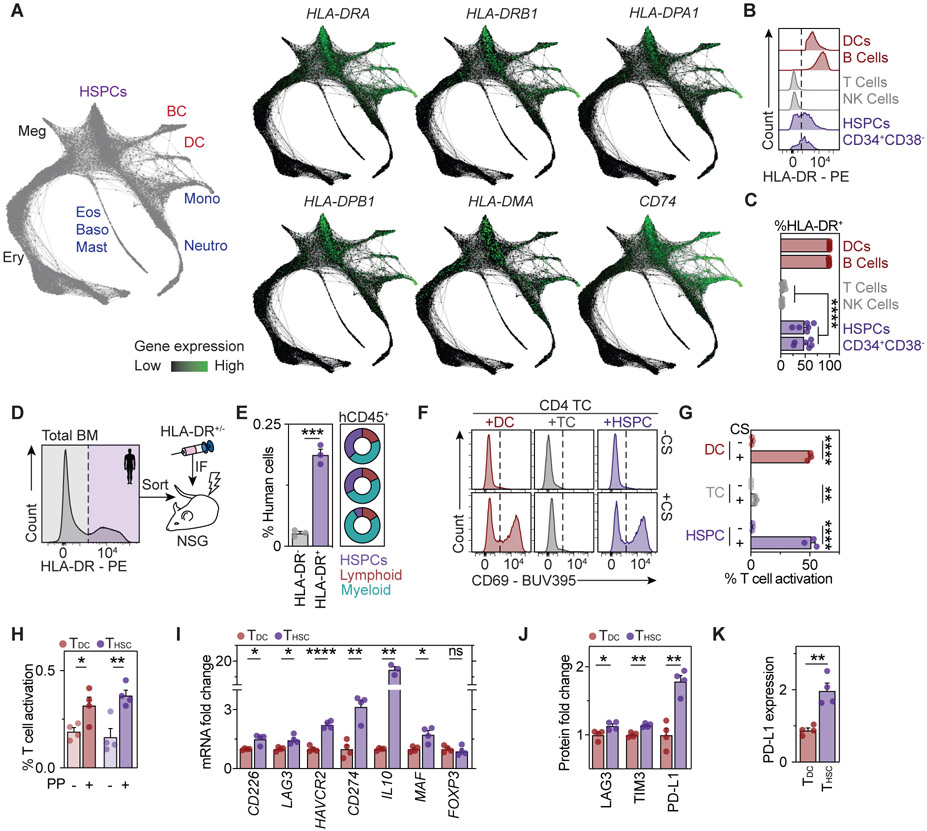
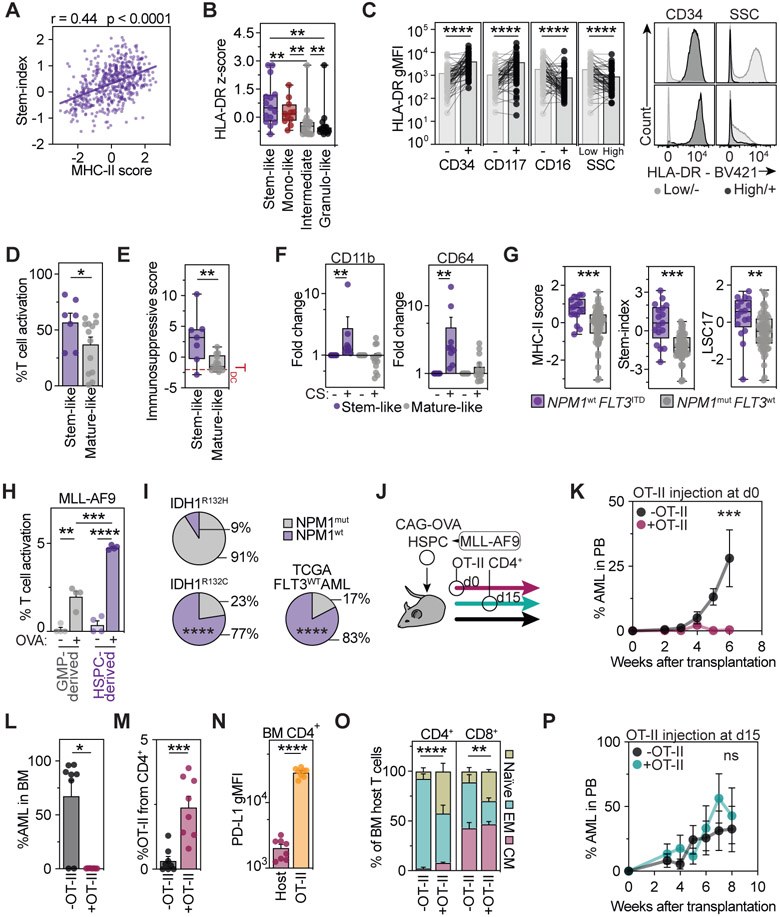
Hematopoietic stem and progenitor cells (HSPCs) are responsible for the production of blood and immune cells. Throughout life, HSPCs acquire oncogenic aberrations that can cause hematological cancers. Although molecular programs maintaining stem cell integrity have been identified, safety mechanisms eliminating malignant HSPCs from the stem cell pool remain poorly characterized. Here, we show that HSPCs constitutively present antigens via major histocompatibility complex class II. The presentation of immunogenic antigens, as occurring during malignant transformation, triggers bidirectional interactions between HSPCs and antigen-specific CD4+ T cells, causing stem cell proliferation, differentiation, and specific exhaustion of aberrant HSPCs. This immunosurveillance mechanism effectively eliminates transformed HSPCs from the hematopoietic system, thereby preventing leukemia onset. Together, our data reveal a bidirectional interaction between HSPCs and CD4+ T cells, demonstrating that HSPCs are not only passive receivers of immunological signals but also actively engage in adaptive immune responses to safeguard the integrity of the stem cell pool.
Keywords: CD4(+) T cells; acute myeloid leukemia; antigen presentation; hematopoietic stem cells; immune regulation; immunosurveillance; leukemia.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests V.K.K. is a cofounder, has ownership interest, and is on the SAB of Celsius Therapeutics and Tizona Therapeutics.
Figures
Comment in
-
Death by differentiation: CD4+ T cells kick out suspicious stem cells.Cell Stem Cell. 2022 May 5;29(5):655-656. doi: 10.1016/j.stem.2022.04.013. Cell Stem Cell. 2022. PMID: 35523132
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
