

Efficient recall of Omicron-reactive B cell memory after a third dose of SARS-CoV-2 mRNA vaccine
- PMID: 35523182
- PMCID: PMC8989683
- DOI: 10.1016/j.cell.2022.04.009
Efficient recall of Omicron-reactive B cell memory after a third dose of SARS-CoV-2 mRNA vaccine
Abstract
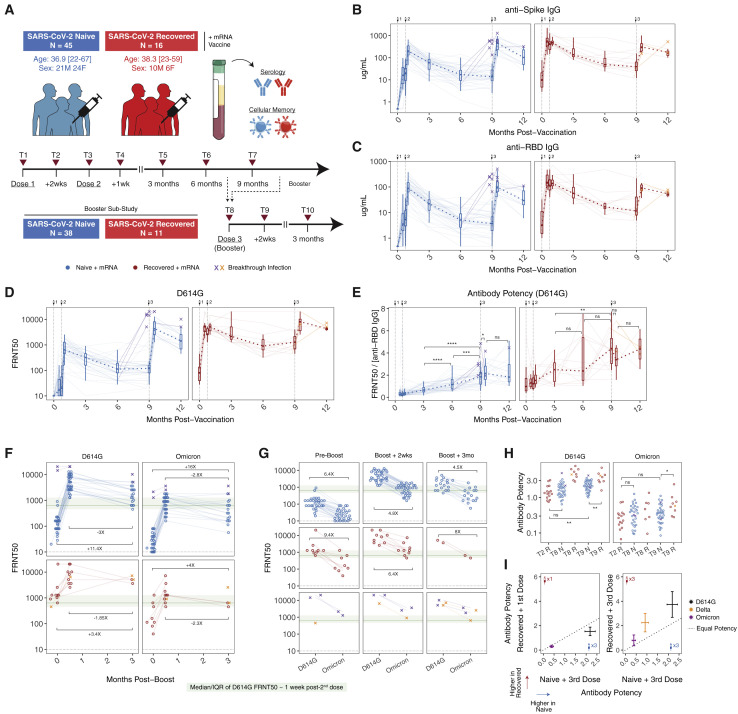
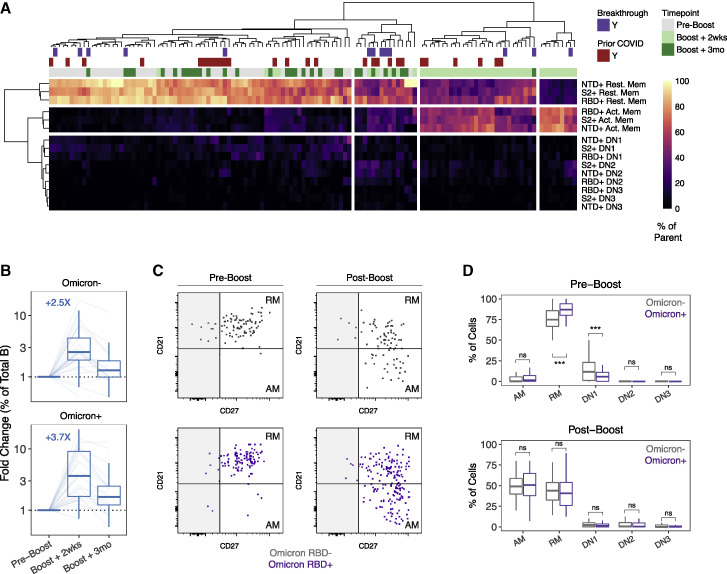
We examined antibody and memory B cell responses longitudinally for ∼9-10 months after primary 2-dose SARS-CoV-2 mRNA vaccination and 3 months after a 3rd dose. Antibody decay stabilized between 6 and 9 months, and antibody quality continued to improve for at least 9 months after 2-dose vaccination. Spike- and RBD-specific memory B cells remained durable over time, and 40%-50% of RBD-specific memory B cells simultaneously bound the Alpha, Beta, Delta, and Omicron variants. Omicron-binding memory B cells were efficiently reactivated by a 3rd dose of wild-type vaccine and correlated with the corresponding increase in neutralizing antibody titers. In contrast, pre-3rd dose antibody titers inversely correlated with the fold-change of antibody boosting, suggesting that high levels of circulating antibodies may limit the added protection afforded by repeat short interval boosting. These data provide insight into the quantity and quality of mRNA-vaccine-induced immunity over time through 3 or more antigen exposures.
Keywords: COVID-19; Omicron; SARS-CoV-2; antibody; booster; immune memory; mRNA; memory B cell; vaccine; variants of concern.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.E.H. has received consultancy fees from Sanofi Pasteur, Lumen, Novavax, and Merck for work unrelated to this study. A.R.G. is a consultant for Relation Therapeutics. E.J.W. is consulting for or is an advisor for Merck, Marengo, Janssen, Related Sciences, Synthekine, and Surface Oncology. E.J.W. is a founder of Surface Oncology, Danger Bio, and Arsenal Biosciences.
Figures







Update of
-
Efficient recall of Omicron-reactive B cell memory after a third dose of SARS-CoV-2 mRNA vaccine.bioRxiv [Preprint]. 2022 Feb 22:2022.02.20.481163. doi: 10.1101/2022.02.20.481163. bioRxiv. 2022. Update in: Cell. 2022 May 26;185(11):1875-1887.e8. doi: 10.1016/j.cell.2022.04.009. PMID: 35233575 Free PMC article. Updated. Preprint.
References
-
- Chatterjee D., Tauzin A., Marchitto L., Gong S.Y., Boutin M., Bourassa C., Beaudoin-Bussières G., Bo Y., Ding S., Laumaea A., et al. SARS-CoV-2 Omicron spike recognition by plasma from individuals receiving BNT162b2 mRNA vaccination with a 16-week interval between doses. Cell Rep. 2022;38:110429. - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- T32 AI007324/AI/NIAID NIH HHS/United States
- T32 AR076951/AR/NIAMS NIH HHS/United States
- R21 AI142638/AI/NIAID NIH HHS/United States
- R01 AI152236/AI/NIAID NIH HHS/United States
- T32 AI070077/AI/NIAID NIH HHS/United States
- T32 CA009140/CA/NCI NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- R38 HL143613/HL/NHLBI NIH HHS/United States
- P30 CA016520/CA/NCI NIH HHS/United States
- 75N93021C00015/AI/NIAID NIH HHS/United States
- T32 AI055400/AI/NIAID NIH HHS/United States
- R01 AI155577/AI/NIAID NIH HHS/United States
- U19 AI149680/AI/NIAID NIH HHS/United States
- T32 HL007843/HL/NHLBI NIH HHS/United States
- P01 AI108545/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
