

Developmental, neurochemical, and behavioral analyses of ErbB4 Cyt-1 knockout mice
- PMID: 35523590
- PMCID: PMC9149141
- DOI: 10.1111/jnc.15612
Developmental, neurochemical, and behavioral analyses of ErbB4 Cyt-1 knockout mice
Abstract
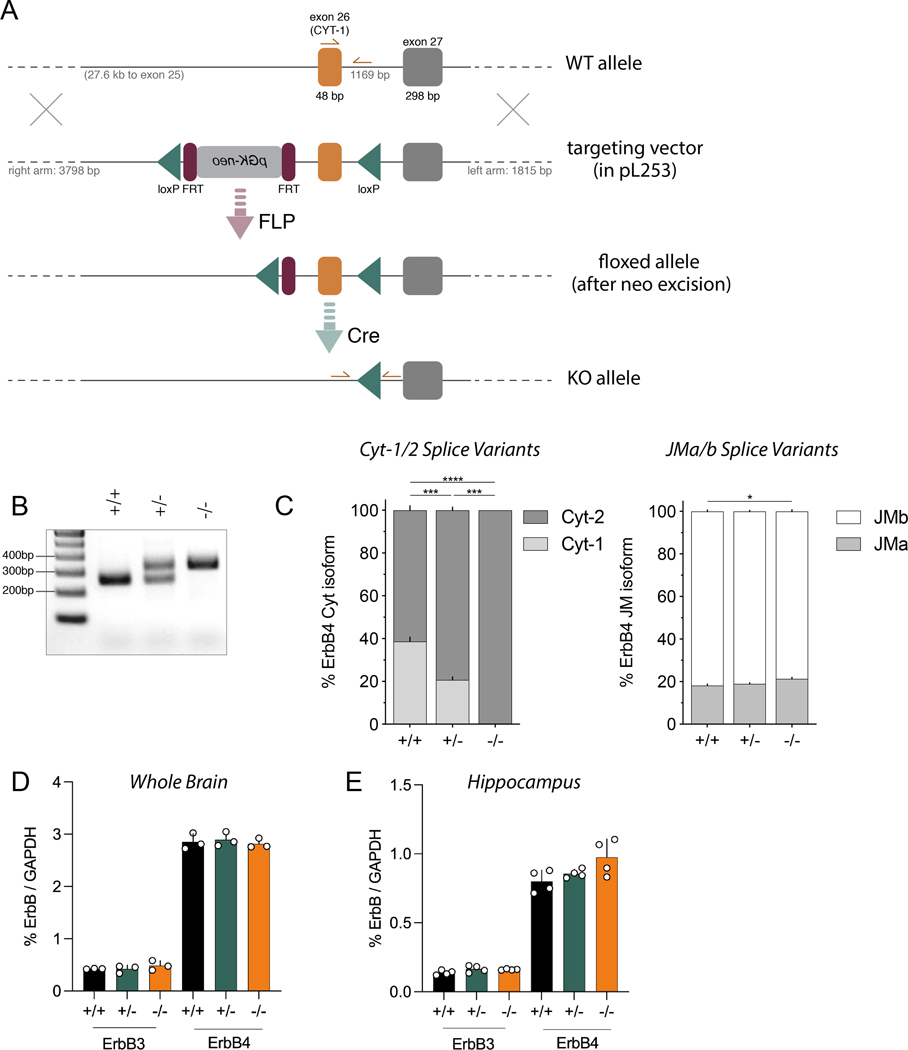
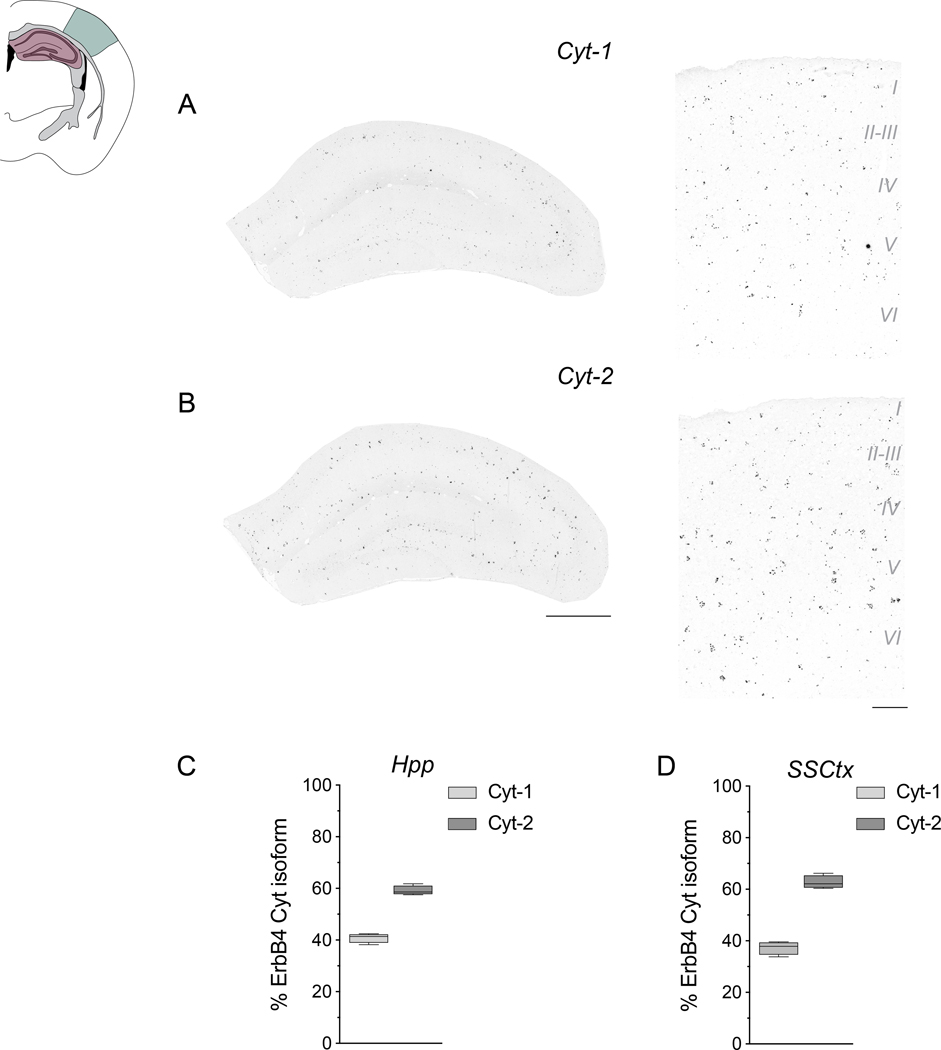
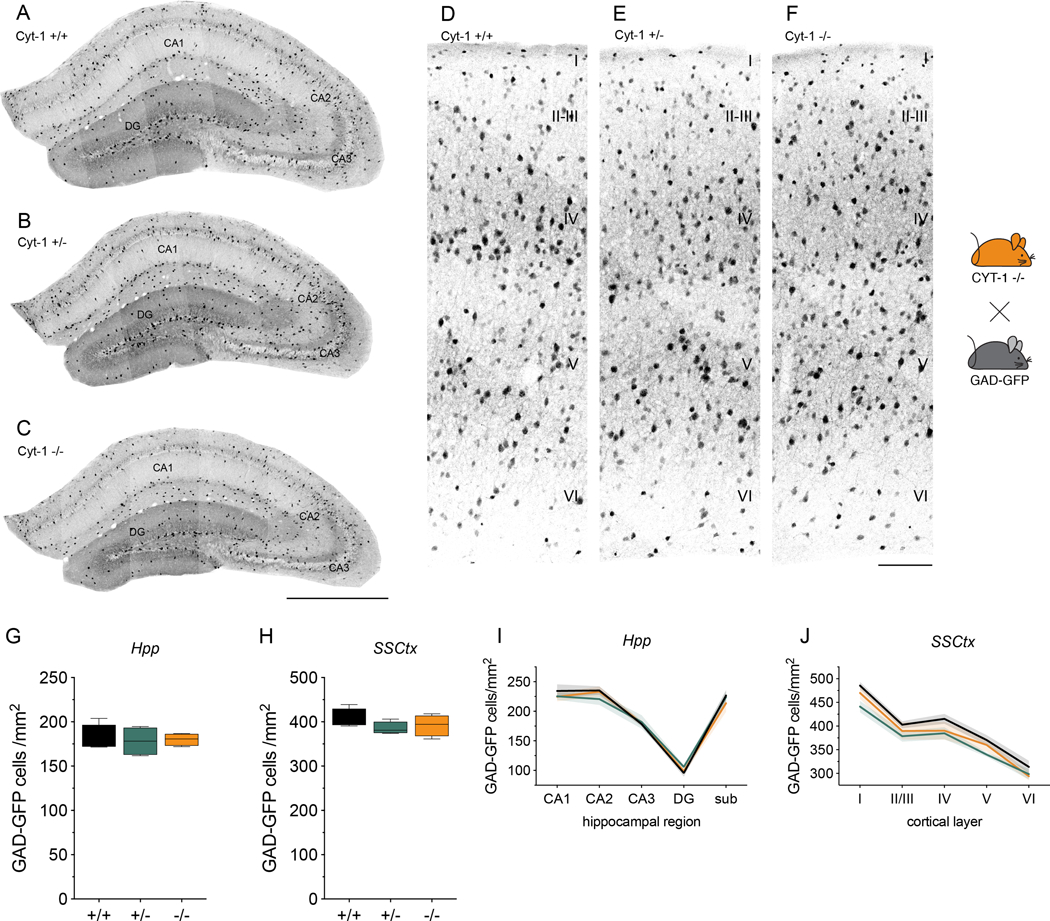
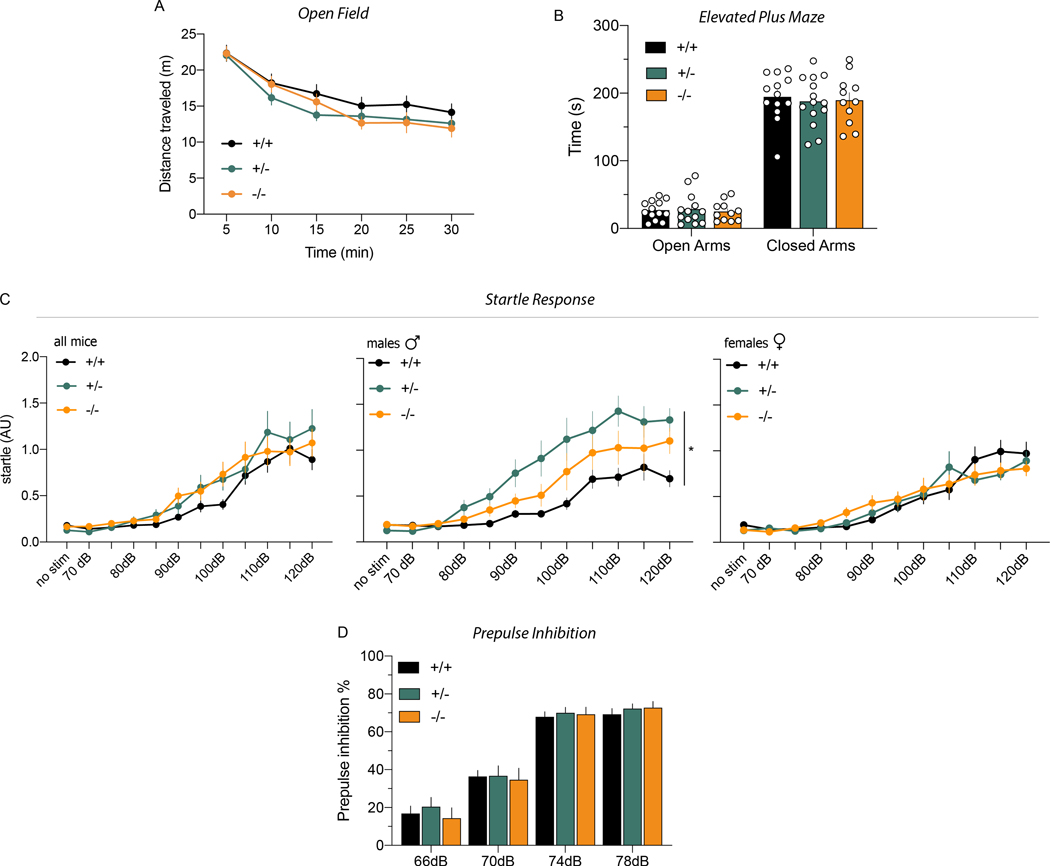
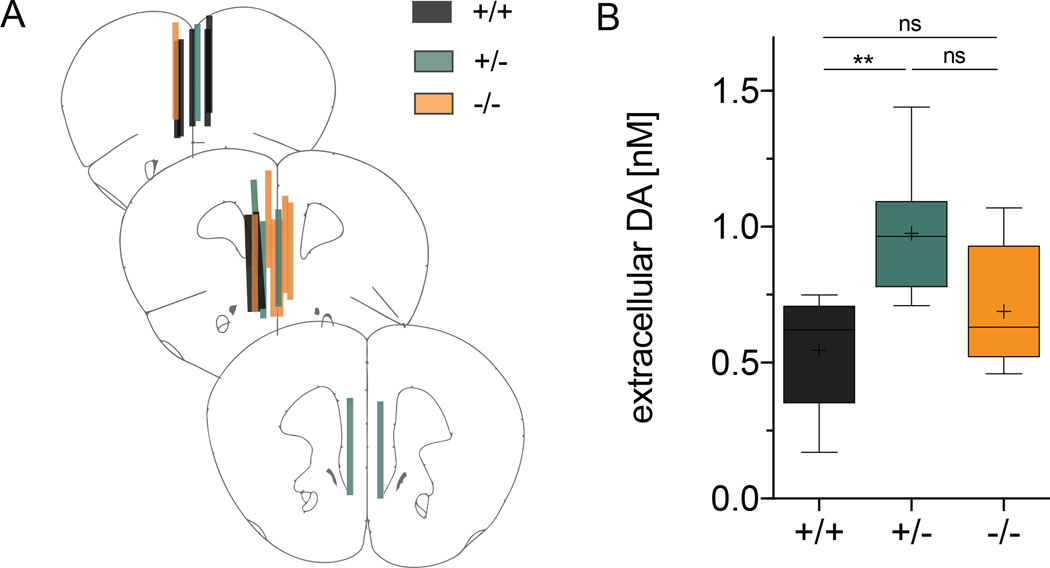
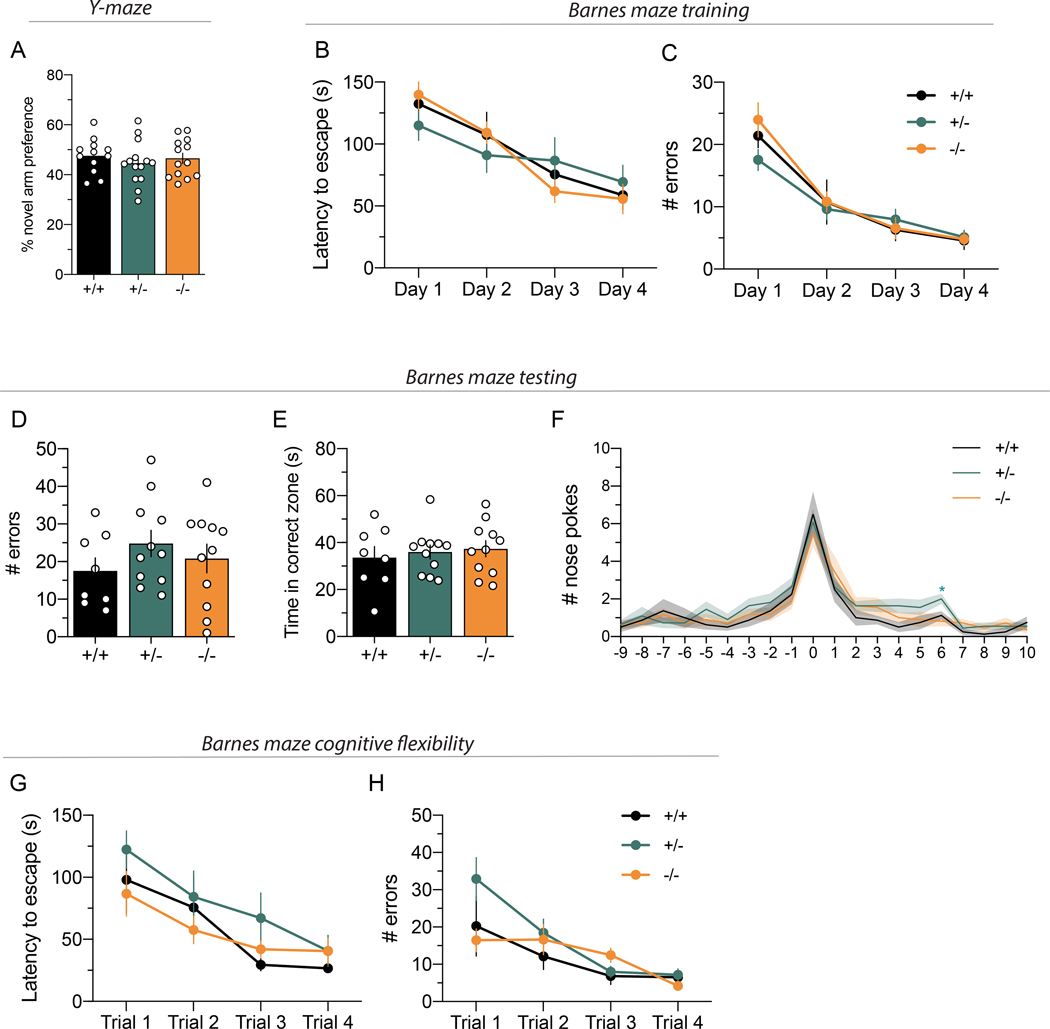
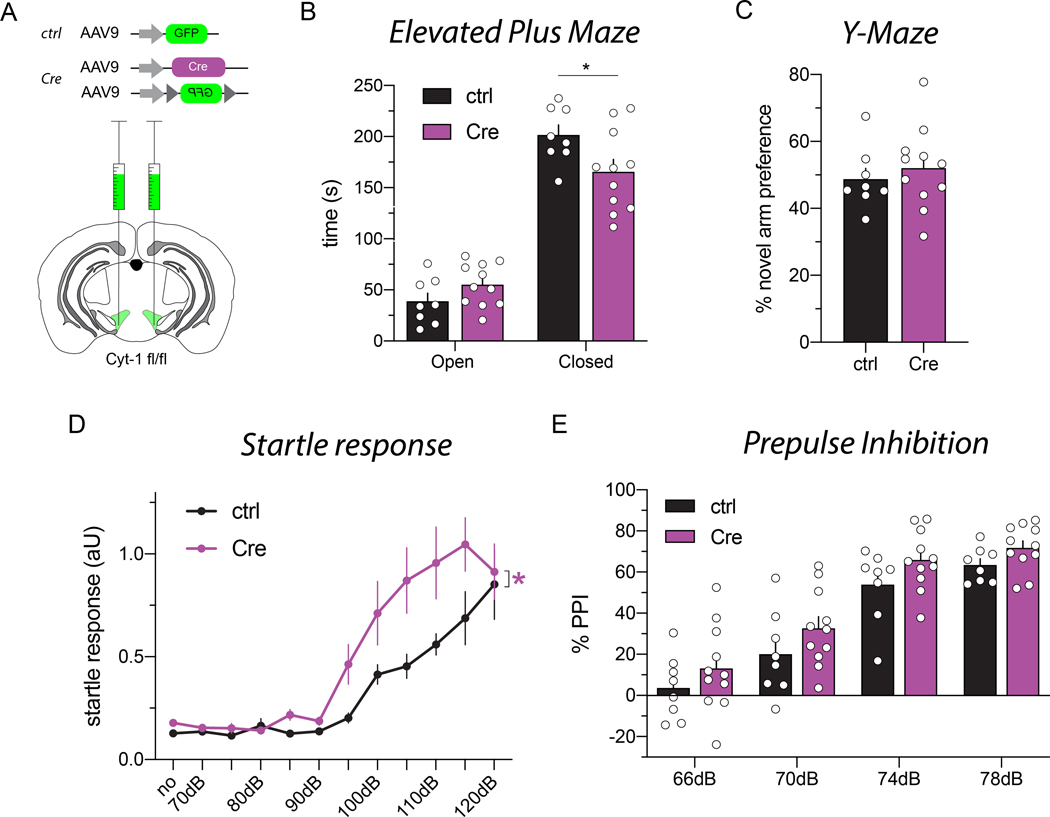
Neuregulins (NRGs) and their cognate neuronal receptor ERBB4, which is expressed in GABAergic and dopaminergic neurons, regulate numerous behaviors in rodents and have been identified as schizophrenia at-risk genes. ErbB4 transcripts are alternatively spliced to generate isoforms that either include (Cyt-1) or exclude (Cyt-2) exon 26, which encodes a cytoplasmic domain that imparts ErbB4 receptors the ability to signal via the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway. Although ErbB4 Cyt-1/2 isoforms have been studied in transfected cultured cells, their functions in vivo remain unknown. Here, we generated ErbB4-floxed (ErbB4-Cyt1fl/fl ) mice to investigate the effects of germline (constitutive) and conditional (acute) deletions of the Cyt-1 exon. Overall receptor mRNA levels remain unchanged in germline ErbB4 Cyt-1 knockouts (Cyt-1 KOs), with all transcripts encoding Cyt-2 variants. In contrast to mice lacking all ErbB4 receptor function, GABAergic interneuron migration and number are unaltered in Cyt-1 KOs. However, basal extracellular dopamine (DA) levels in the medial prefrontal cortex are increased in Cyt-1 heterozygotes. Despite these neurochemical changes, Cyt-1 heterozygous and homozygous mice do not manifest behavioral abnormalities previously reported to be altered in ErbB4 null mice. To address the possibility that Cyt-2 variants compensate for the lack of Cyt-1 during development, we microinjected an adeno-associated virus expressing Cre-recombinase (AAV-Cre) into the DA-rich ventral tegmental area of adult ErbB4-Cyt1fl/fl mice to acutely target exon 26. These conditional Cyt-1 KOs were found to exhibit behavioral abnormalities in the elevated plus maze and startle response, consistent with the idea that late exon 26 ablations may circumvent compensation by Cyt-2 variants. Taken together, our observations indicate that ErbB4 Cyt-1 function in vivo is important for DA balance and behaviors in adults.
© 2022 The Authors. Journal of Neurochemistry published by John Wiley & Sons Ltd on behalf of International Society for Neurochemistry. This article has been contributed to by U.S. Government employees and their work is in the public domain in the USA.
Conflict of interest statement
Conflicts of Interest
Andrés Buonanno is a senior editor for the Journal of Neurochemistry. No additional conflicts of interest to report.
Figures
References
-
- Abe Y, Namba H, Zheng Y. and Nawa H. (2009) In situ hybridization reveals developmental regulation of ErbB1–4 mRNA expression in mouse midbrain: implication of ErbB receptors for dopaminergic neurons. Neuroscience 161, 95–110. - PubMed
-
- Buonanno A. and Fischbach GD (2001) Neuregulin and ErbB receptor signaling pathways in the nervous system. Curr Opin Neurobiol 11, 287–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
