

Regulation of A-to-I RNA editing and stop codon recoding to control selenoprotein expression during skeletal myogenesis
- PMID: 35523818
- PMCID: PMC9076623
- DOI: 10.1038/s41467-022-30181-2
Regulation of A-to-I RNA editing and stop codon recoding to control selenoprotein expression during skeletal myogenesis
Abstract
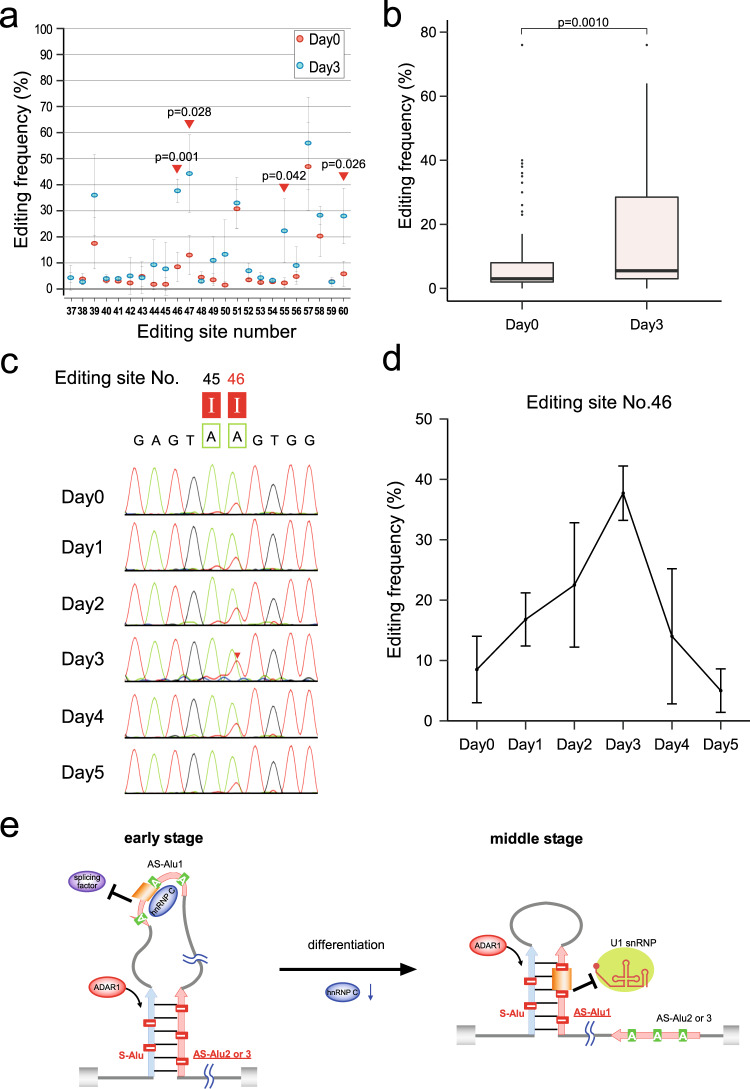
Selenoprotein N (SELENON), a selenocysteine (Sec)-containing protein with high reductive activity, maintains redox homeostasis, thereby contributing to skeletal muscle differentiation and function. Loss-of-function mutations in SELENON cause severe neuromuscular disorders. In the early-to-middle stage of myoblast differentiation, SELENON maintains redox homeostasis and modulates endoplasmic reticulum (ER) Ca2+ concentration, resulting in a gradual reduction from the middle-to-late stages due to unknown mechanisms. The present study describes post-transcriptional mechanisms that regulate SELENON expression during myoblast differentiation. Part of an Alu element in the second intron of SELENON pre-mRNA is frequently exonized during splicing, resulting in an aberrant mRNA that is degraded by nonsense-mediated mRNA decay (NMD). In the middle stage of myoblast differentiation, ADAR1-mediated A-to-I RNA editing occurs in the U1 snRNA binding site at 5' splice site, preventing Alu exonization and producing mature mRNA. In the middle-to-late stage of myoblast differentiation, the level of Sec-charged tRNASec decreases due to downregulation of essential recoding factors for Sec insertion, thereby generating a premature termination codon in SELENON mRNA, which is targeted by NMD.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
A Versatile Strategy to Reduce UGA-Selenocysteine Recoding Efficiency of the Ribosome Using CRISPR-Cas9-Viral-Like-Particles Targeting Selenocysteine-tRNA[Ser]Sec Gene.Cells. 2019 Jun 11;8(6):574. doi: 10.3390/cells8060574. Cells. 2019. PMID: 31212706 Free PMC article.
-
Partitioning between recoding and termination at a stop codon-selenocysteine insertion sequence.Nucleic Acids Res. 2015 Jul 27;43(13):6426-38. doi: 10.1093/nar/gkv558. Epub 2015 Jun 3. Nucleic Acids Res. 2015. PMID: 26040702 Free PMC article.
-
Modeling and gene knockdown to assess the contribution of nonsense-mediated decay, premature termination, and selenocysteine insertion to the selenoprotein hierarchy.RNA. 2016 Jul;22(7):1076-84. doi: 10.1261/rna.055749.115. Epub 2016 May 20. RNA. 2016. PMID: 27208313 Free PMC article.
-
Update on selenoprotein biosynthesis.Antioxid Redox Signal. 2015 Oct 1;23(10):775-94. doi: 10.1089/ars.2015.6391. Epub 2015 Aug 25. Antioxid Redox Signal. 2015. PMID: 26154496 Review.
-
The unique tRNASec and its role in selenocysteine biosynthesis.Amino Acids. 2018 Sep;50(9):1145-1167. doi: 10.1007/s00726-018-2595-6. Epub 2018 Jun 12. Amino Acids. 2018. PMID: 29948343 Review.
Cited by
-
A Wonderful Journey: The Diverse Roles of Adenosine Deaminase Action on RNA 1 (ADAR1) in Central Nervous System Diseases.CNS Neurosci Ther. 2025 Jan;31(1):e70208. doi: 10.1111/cns.70208. CNS Neurosci Ther. 2025. PMID: 39753993 Free PMC article. Review.
-
Alterations in RNA editing in skeletal muscle following exercise training in individuals with Parkinson's disease.PLoS One. 2023 Dec 22;18(12):e0287078. doi: 10.1371/journal.pone.0287078. eCollection 2023. PLoS One. 2023. PMID: 38134032 Free PMC article.
-
ADARs: pleiotropy in function, versatility in application.Nucleic Acids Res. 2025 Jul 8;53(13):gkaf672. doi: 10.1093/nar/gkaf672. Nucleic Acids Res. 2025. PMID: 40671518 Free PMC article. Review.
-
A comprehensive atlas of pig RNA editome across 23 tissues reveals RNA editing affecting interaction mRNA-miRNAs.G3 (Bethesda). 2024 Oct 7;14(10):jkae178. doi: 10.1093/g3journal/jkae178. G3 (Bethesda). 2024. PMID: 39090686 Free PMC article.
-
The role of dsRNA A-to-I editing catalyzed by ADAR family enzymes in the pathogeneses.RNA Biol. 2024 Jan;21(1):52-69. doi: 10.1080/15476286.2024.2414156. Epub 2024 Oct 24. RNA Biol. 2024. PMID: 39449182 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
