

The MFN1 and MFN2 mitofusins promote clustering between mitochondria and peroxisomes
- PMID: 35523862
- PMCID: PMC9076876
- DOI: 10.1038/s42003-022-03377-x
The MFN1 and MFN2 mitofusins promote clustering between mitochondria and peroxisomes
Abstract
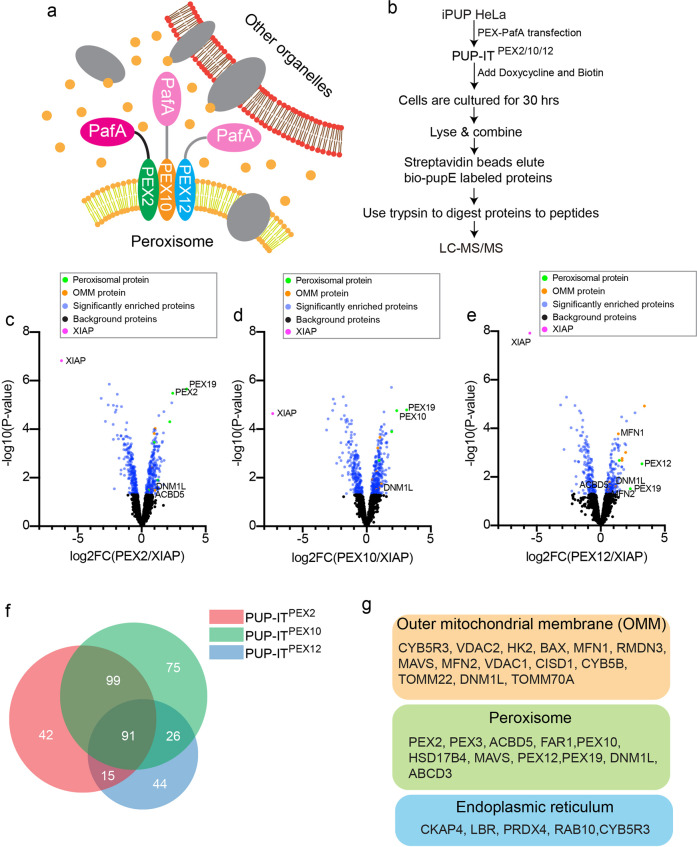
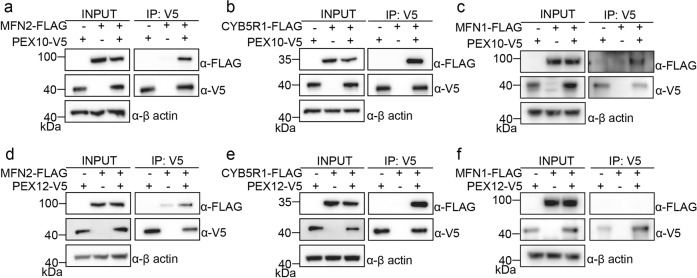
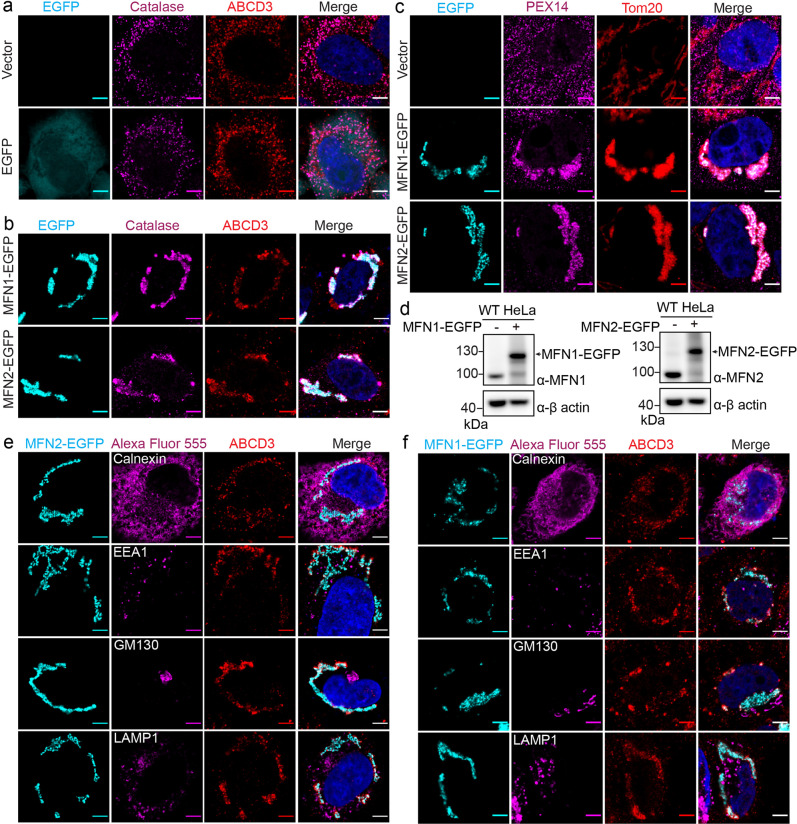
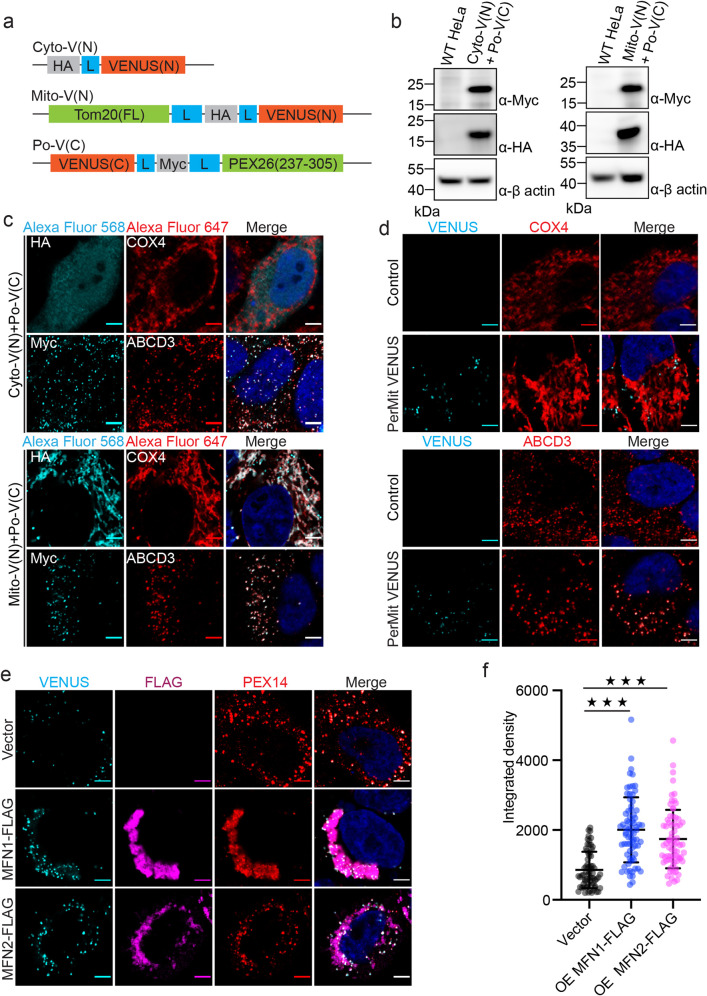
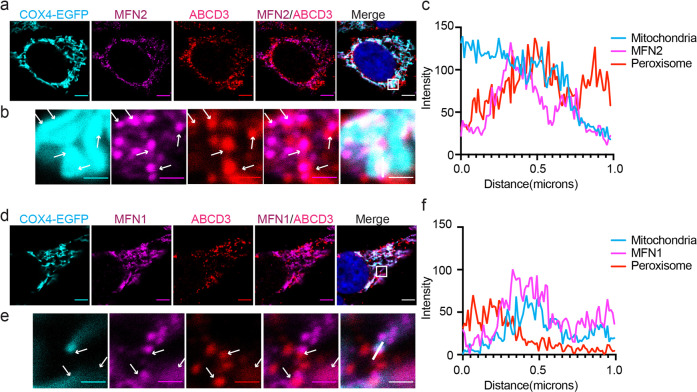
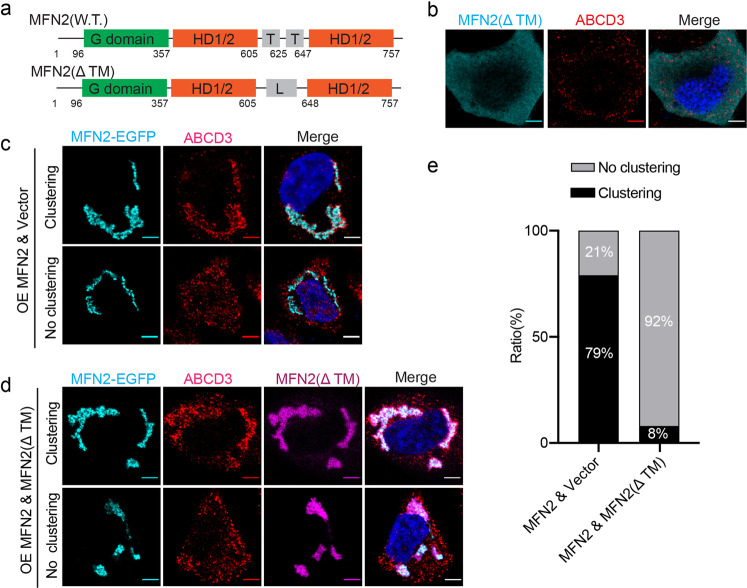
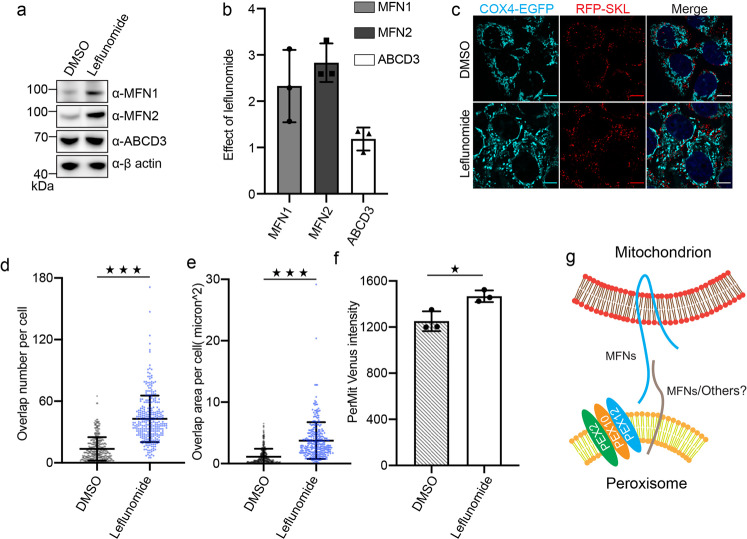
Mitochondria and peroxisomes are two types of functionally close-related organelles, and both play essential roles in lipid and ROS metabolism. However, how they physically interact with each other is not well understood. In this study, we apply the proximity labeling method with peroxisomal proteins and report that mitochondrial protein mitofusins (MFNs) are in proximity to peroxisomes. Overexpression of MFNs induces not only the mitochondria clustering but also the co-clustering of peroxisomes. We also report the enrichment of MFNs at the mitochondria-peroxisome interface. Induced mitofusin expression gives rise to more mitochondria-peroxisome contacting sites. Furthermore, the tethering of peroxisomes to mitochondria can be inhibited by the expression of a truncated MFN2, which lacks the transmembrane region. Collectively, our study suggests MFNs as regulators for mitochondria-peroxisome contacts. Our findings are essential for future studies of inter-organelle metabolism regulation and signaling, and may help understand the pathogenesis of mitofusin dysfunction-related disease.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests. Min Zhuang is an Editorial Board Member for
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
