

Walking strides direct rapid and flexible recruitment of visual circuits for course control in Drosophila
- PMID: 35525243
- PMCID: PMC9275417
- DOI: 10.1016/j.neuron.2022.04.008
Walking strides direct rapid and flexible recruitment of visual circuits for course control in Drosophila
Abstract
Flexible mapping between activity in sensory systems and movement parameters is a hallmark of motor control. This flexibility depends on the continuous comparison of short-term postural dynamics and the longer-term goals of an animal, thereby necessitating neural mechanisms that can operate across multiple timescales. To understand how such body-brain interactions emerge across timescales to control movement, we performed whole-cell patch recordings from visual neurons involved in course control in Drosophila. We show that the activity of leg mechanosensory cells, propagating via specific ascending neurons, is critical for stride-by-stride steering adjustments driven by the visual circuit, and, at longer timescales, it provides information about the moving body's state to flexibly recruit the visual circuit for course control. Thus, our findings demonstrate the presence of an elegant stride-based mechanism operating at multiple timescales for context-dependent course control. We propose that this mechanism functions as a general basis for the adaptive control of locomotion.
Keywords: Drosophila; ascending neurons; locomotion control; motor context; multi-timescale processing; steering; stride cycle; visuomotor integration.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
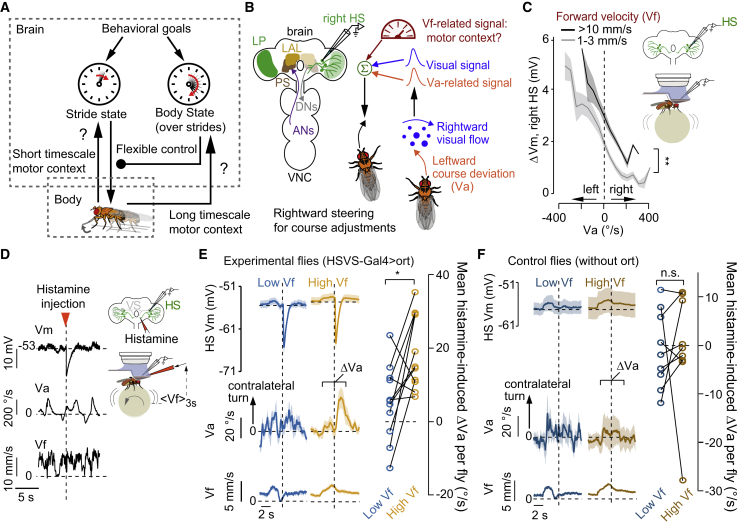
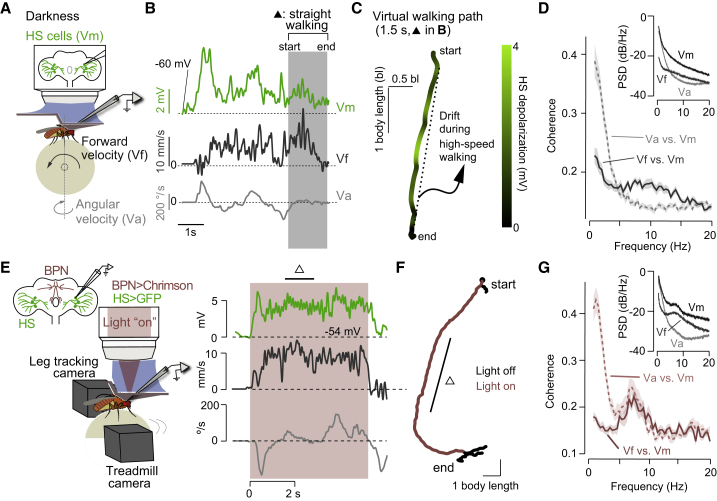
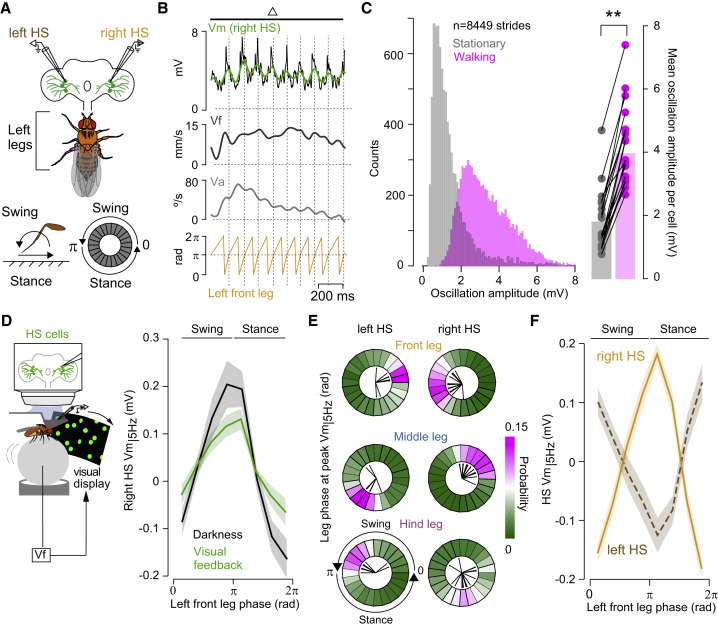
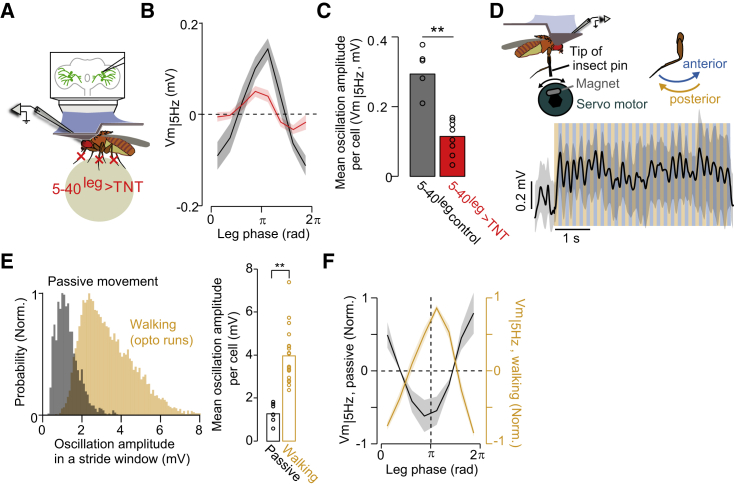
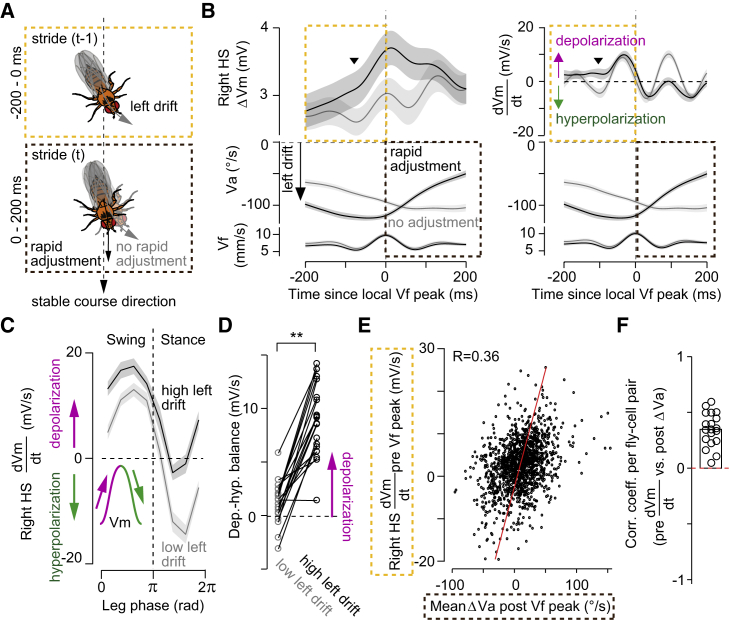
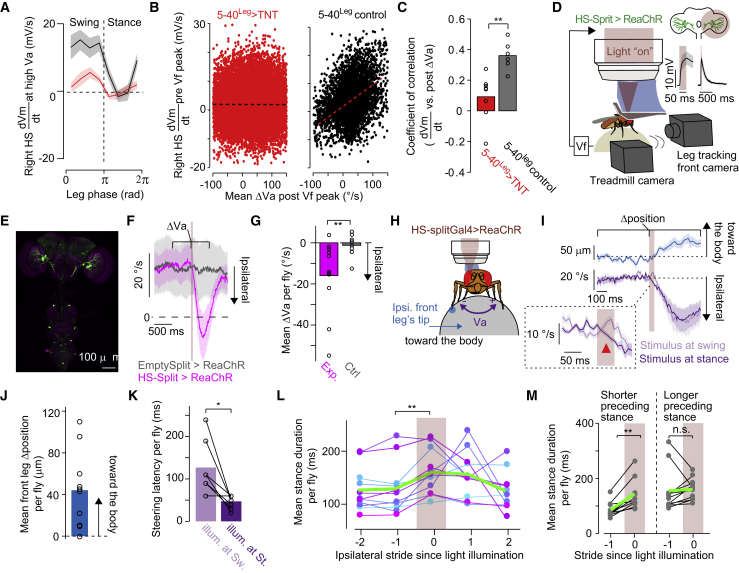
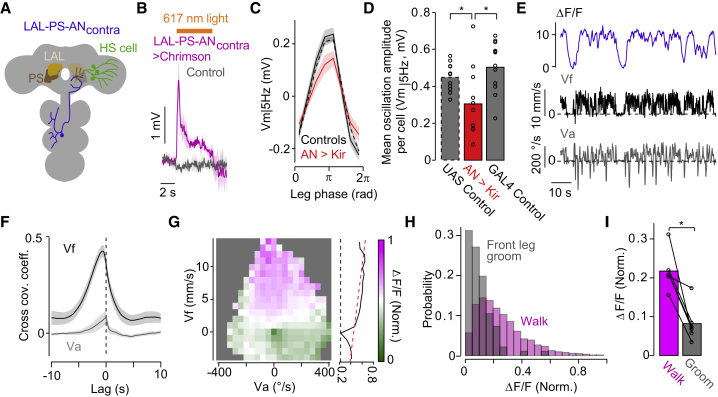
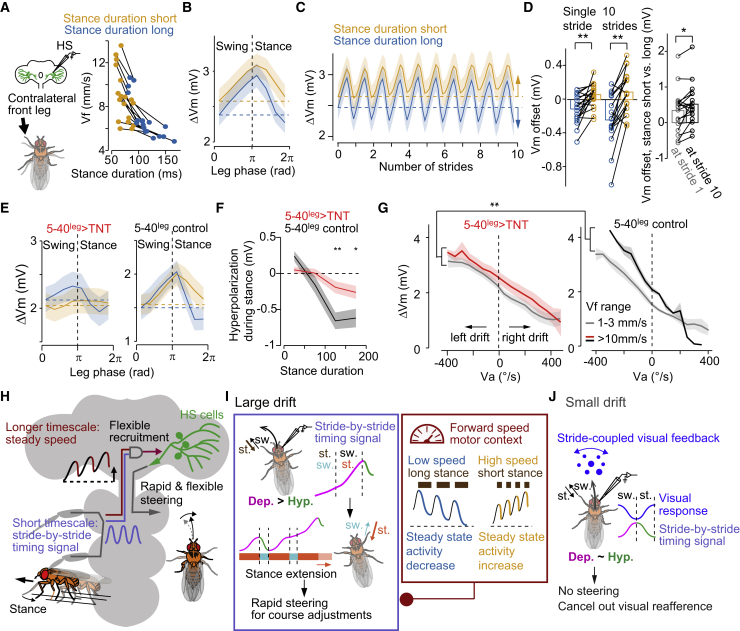
References
-
- Bargmann C.I. Beyond the connectome: how neuromodulators shape neural circuits. BioEssays. 2012;34:458–465. - PubMed
-
- Bässler U., Büschges A. Pattern generation for stick insect walking movements—multisensory control of a locomotor program. Brain Res. Brain Res. Rev. 1998;27:65–88. - PubMed
-
- Bernstein N. Pergamon Press; 1967. The Coordination and Regulation of Movements.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
