

Recurrent inversion polymorphisms in humans associate with genetic instability and genomic disorders
- PMID: 35525246
- PMCID: PMC9563103
- DOI: 10.1016/j.cell.2022.04.017
Recurrent inversion polymorphisms in humans associate with genetic instability and genomic disorders
Abstract
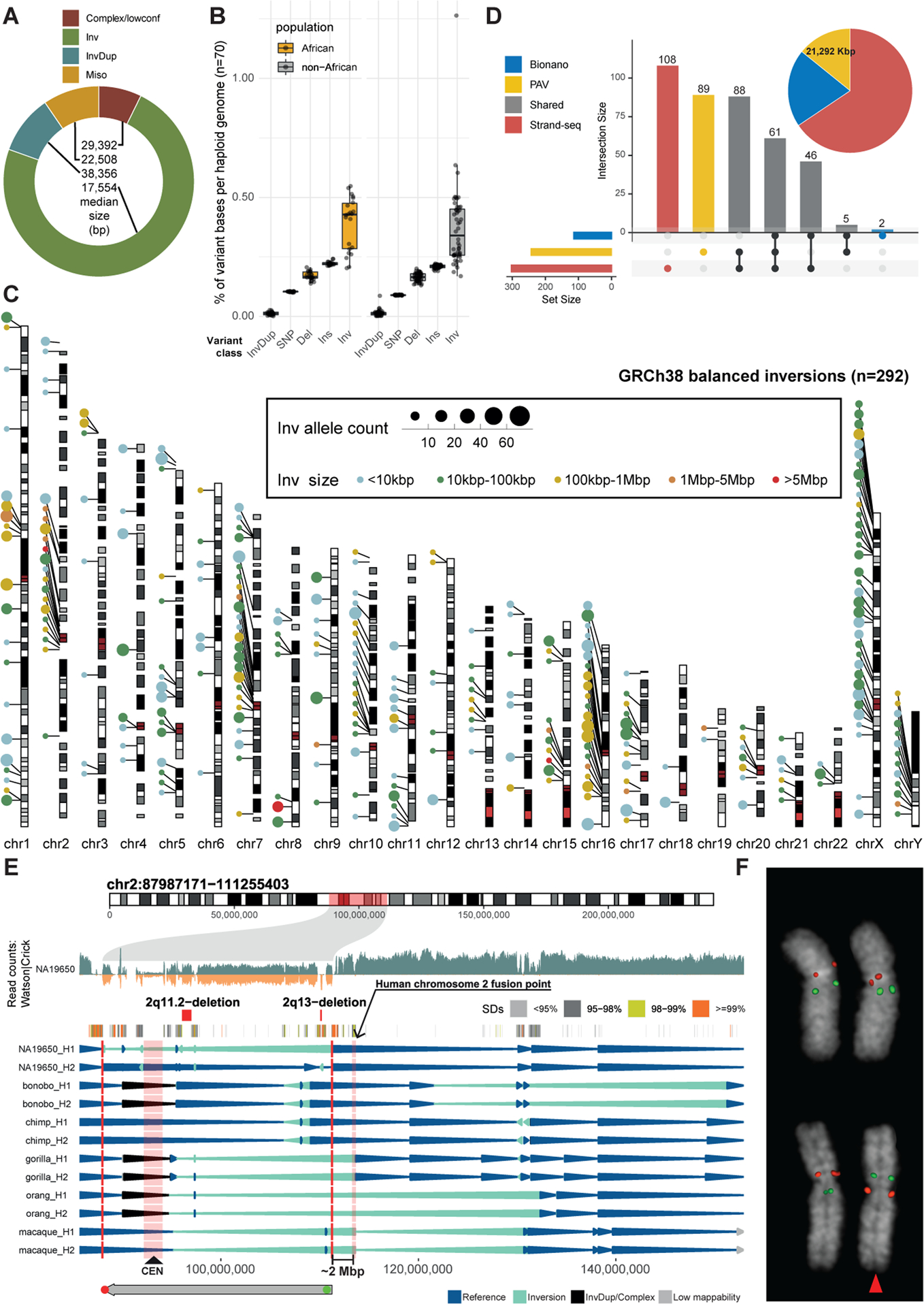
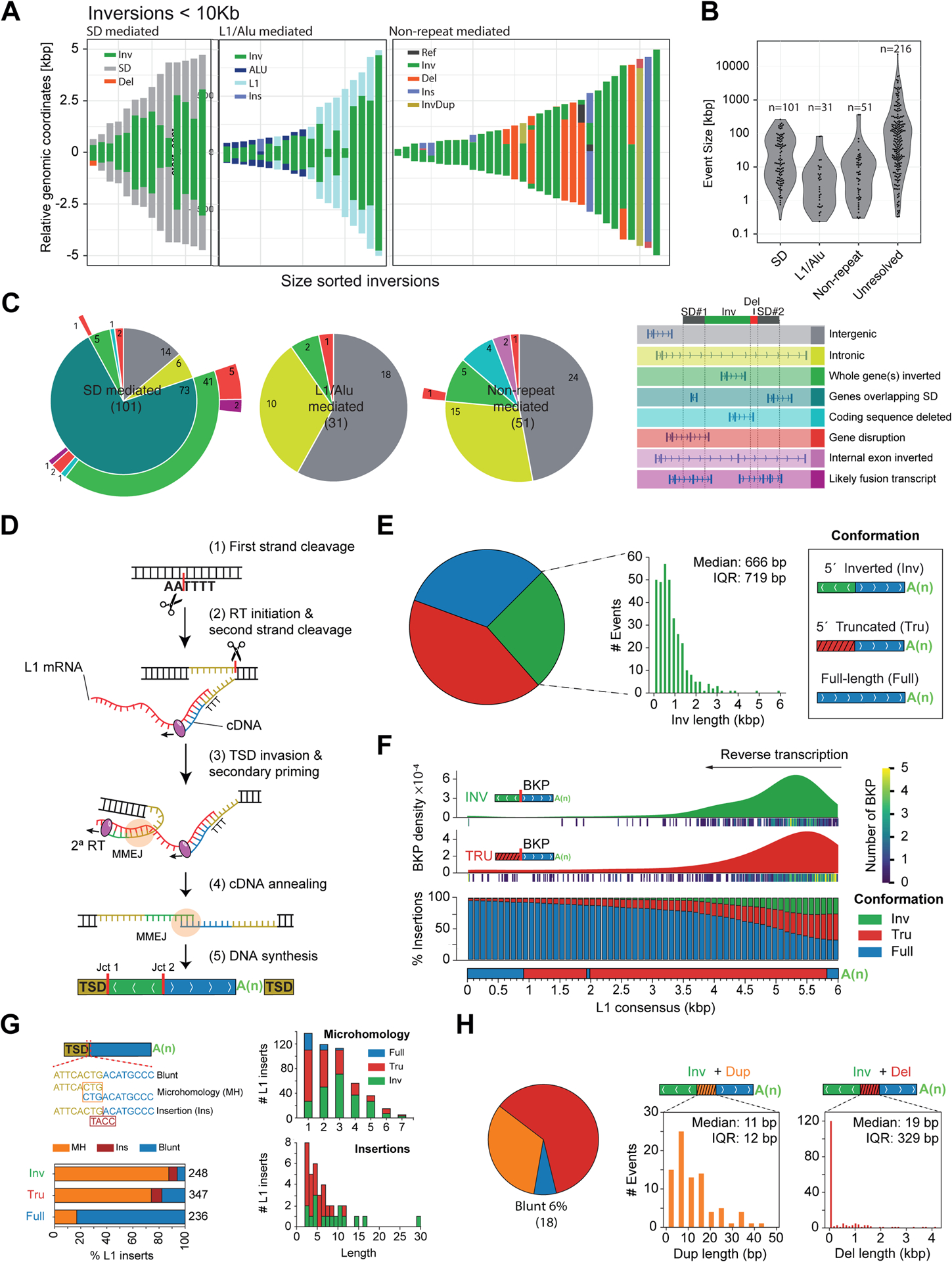
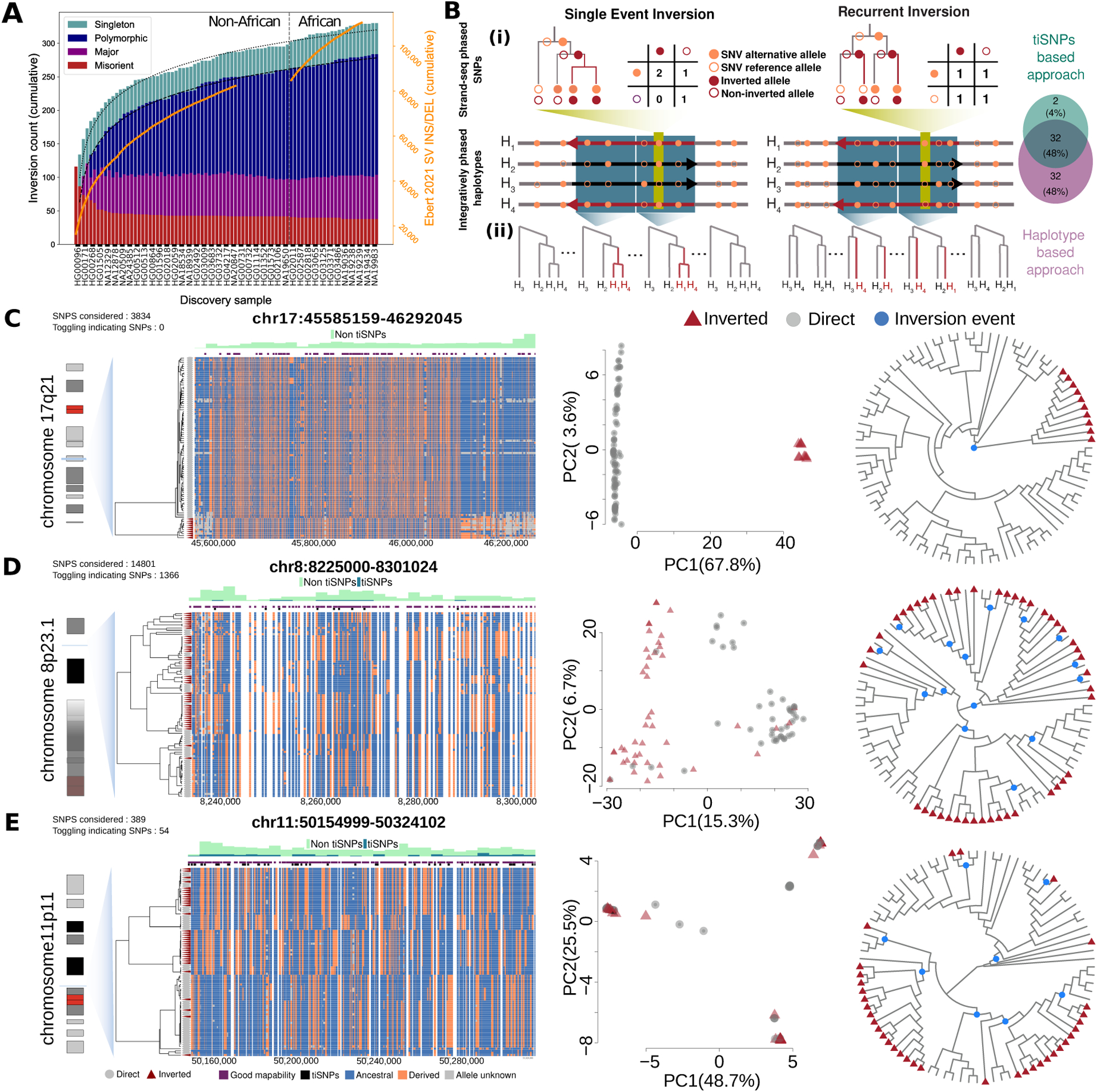
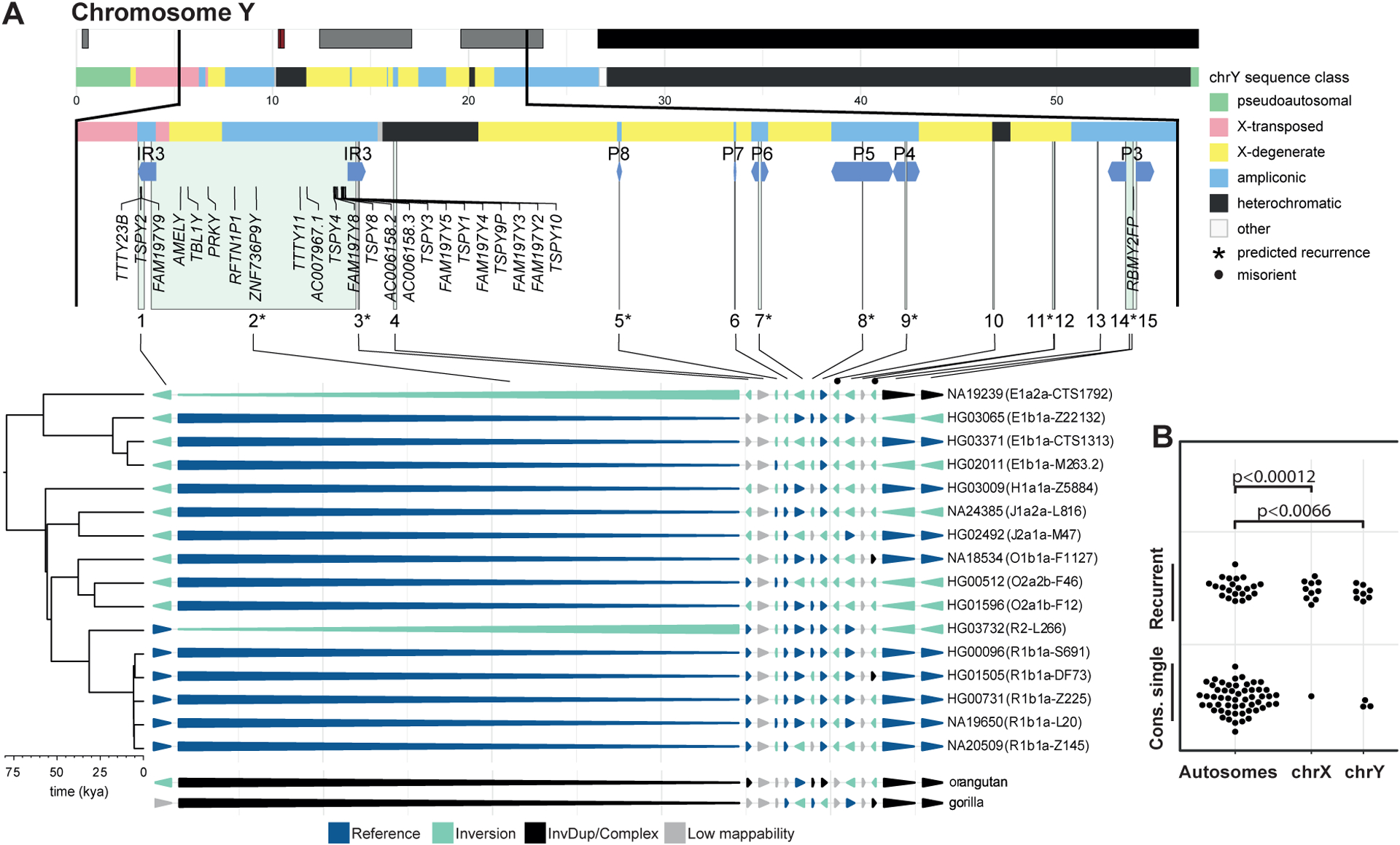
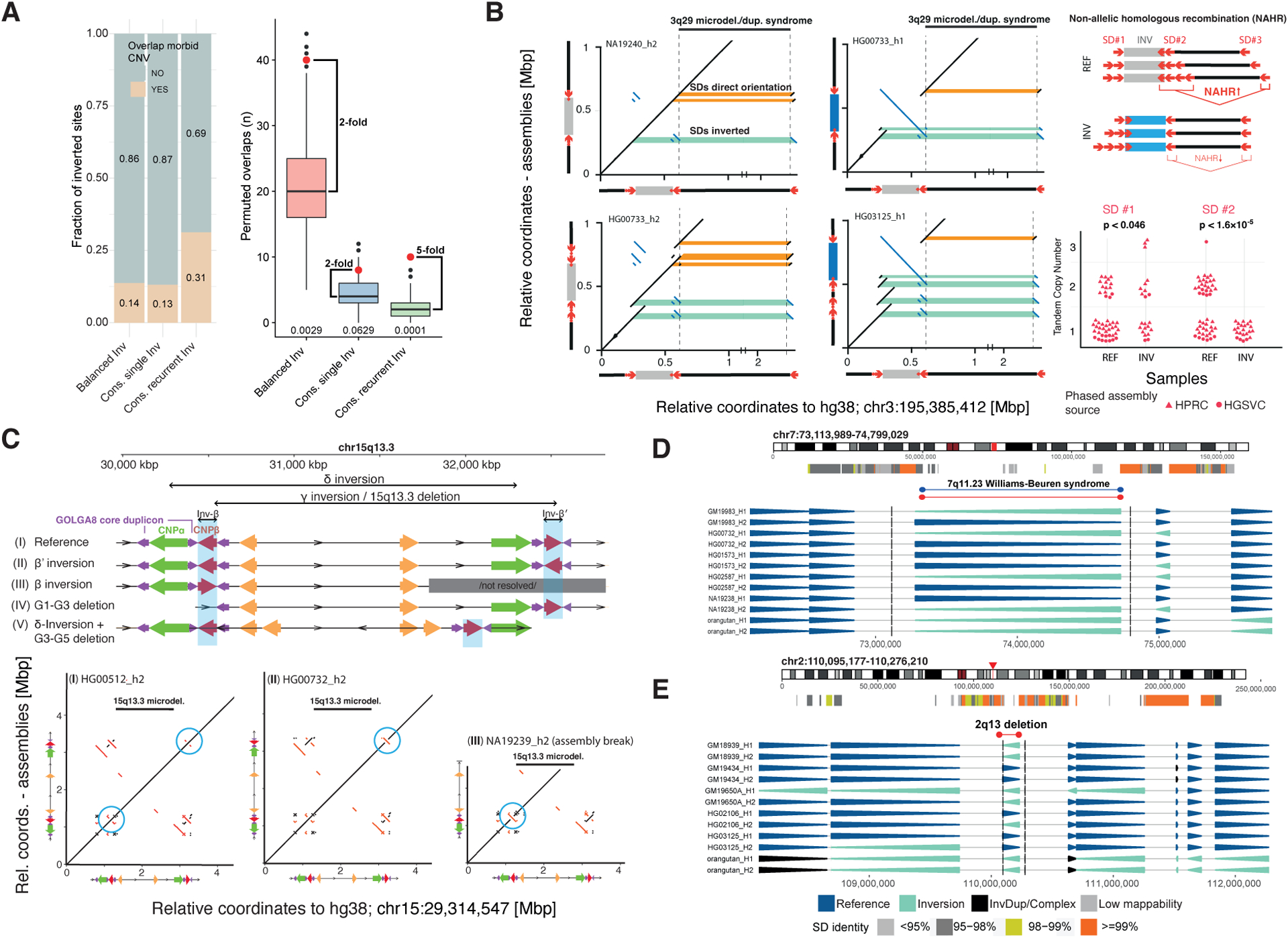
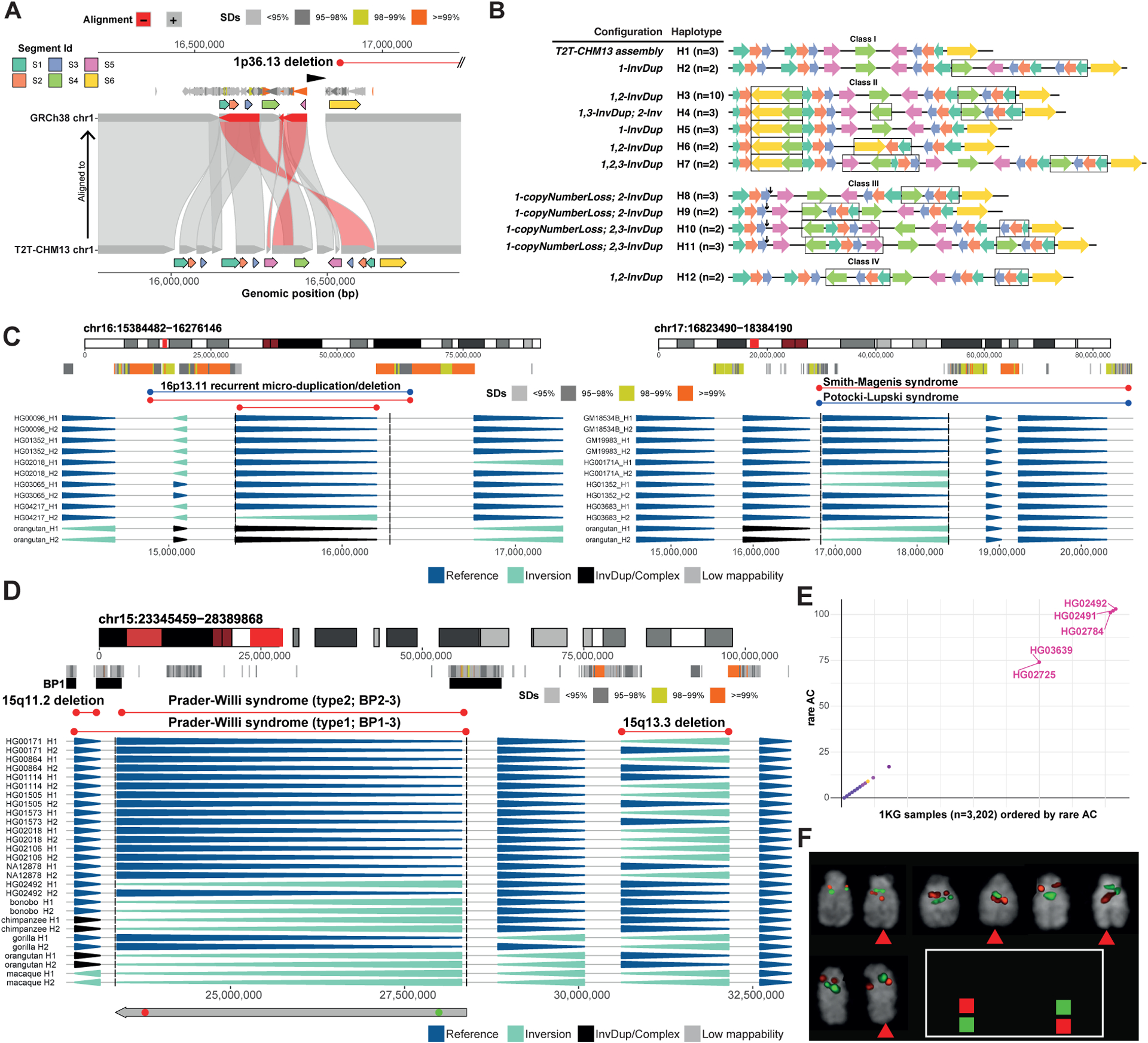
Unlike copy number variants (CNVs), inversions remain an underexplored genetic variation class. By integrating multiple genomic technologies, we discover 729 inversions in 41 human genomes. Approximately 85% of inversions <2 kbp form by twin-priming during L1 retrotransposition; 80% of the larger inversions are balanced and affect twice as many nucleotides as CNVs. Balanced inversions show an excess of common variants, and 72% are flanked by segmental duplications (SDs) or retrotransposons. Since flanking repeats promote non-allelic homologous recombination, we developed complementary approaches to identify recurrent inversion formation. We describe 40 recurrent inversions encompassing 0.6% of the genome, showing inversion rates up to 2.7 × 10-4 per locus per generation. Recurrent inversions exhibit a sex-chromosomal bias and co-localize with genomic disorder critical regions. We propose that inversion recurrence results in an elevated number of heterozygous carriers and structural SD diversity, which increases mutability in the population and predisposes specific haplotypes to disease-causing CNVs.
Keywords: L1 mobile element; genomic disorder; genomic instability; genomic structural variation; human genetic variation; inversion; pathogenic CNV; recurrent mutation; retrotransposon.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.E.E. is a scientific advisory board (SAB) member of Variant Bio. C.L. is an SAB member of Nabsys. The following authors have previously disclosed a patent application (no. EP19169090) relevant to Strand-seq: A.D.S., J.O.K., T.M., and D.P.; the other authors declare no competing interests.
Figures
References
-
- Aagaard Nolting L, Brasch-Andersen C, Cox H, Kanani F, Parker M, Fry AE, Loddo S, Novelli A, Dentici ML, Joss S, et al. (2020). A new 1p36.13-1p36.12 microdeletion syndrome characterized by learning disability, behavioral abnormalities, and ptosis. Clin. Genet 97, 927–932. - PubMed
-
- Aguado C, Gayà-Vidal M, Villatoro S, Oliva M, Izquierdo D, Giner-Delgado C, Montalvo V, García-González J, Martínez-Fundichely A, Capilla L, et al. (2014). Validation and genotyping of multiple human polymorphic inversions mediated by inverted repeats reveals a high degree of recurrence. PLoS Genet. 10, e1004208. - PMC - PubMed
-
- Anantharaman TS, Mysore V, and Mishra B (2004). Fast and cheap genome wide haplotype construction via optical mapping. Biocomputing 2005. - PubMed
Publication types
MeSH terms
Grants and funding
- K99 HG011041/HG/NHGRI NIH HHS/United States
- U01 HG010961/HG/NHGRI NIH HHS/United States
- R21 CA259309/CA/NCI NIH HHS/United States
- R35 GM138212/GM/NIGMS NIH HHS/United States
- U01 HG010973/HG/NHGRI NIH HHS/United States
- P30 CA034196/CA/NCI NIH HHS/United States
- UM1 HG008901/HG/NHGRI NIH HHS/United States
- R01 HG010169/HG/NHGRI NIH HHS/United States
- U01 HG010963/HG/NHGRI NIH HHS/United States
- R01 CA261934/CA/NCI NIH HHS/United States
- U01 HG010971/HG/NHGRI NIH HHS/United States
- U24 HG007497/HG/NHGRI NIH HHS/United States
- R01 HG002385/HG/NHGRI NIH HHS/United States
- R35 GM133600/GM/NIGMS NIH HHS/United States
- U41 HG010972/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
