

Lymph node colonization induces tumor-immune tolerance to promote distant metastasis
- PMID: 35525247
- PMCID: PMC9149144
- DOI: 10.1016/j.cell.2022.04.019
Lymph node colonization induces tumor-immune tolerance to promote distant metastasis
Abstract
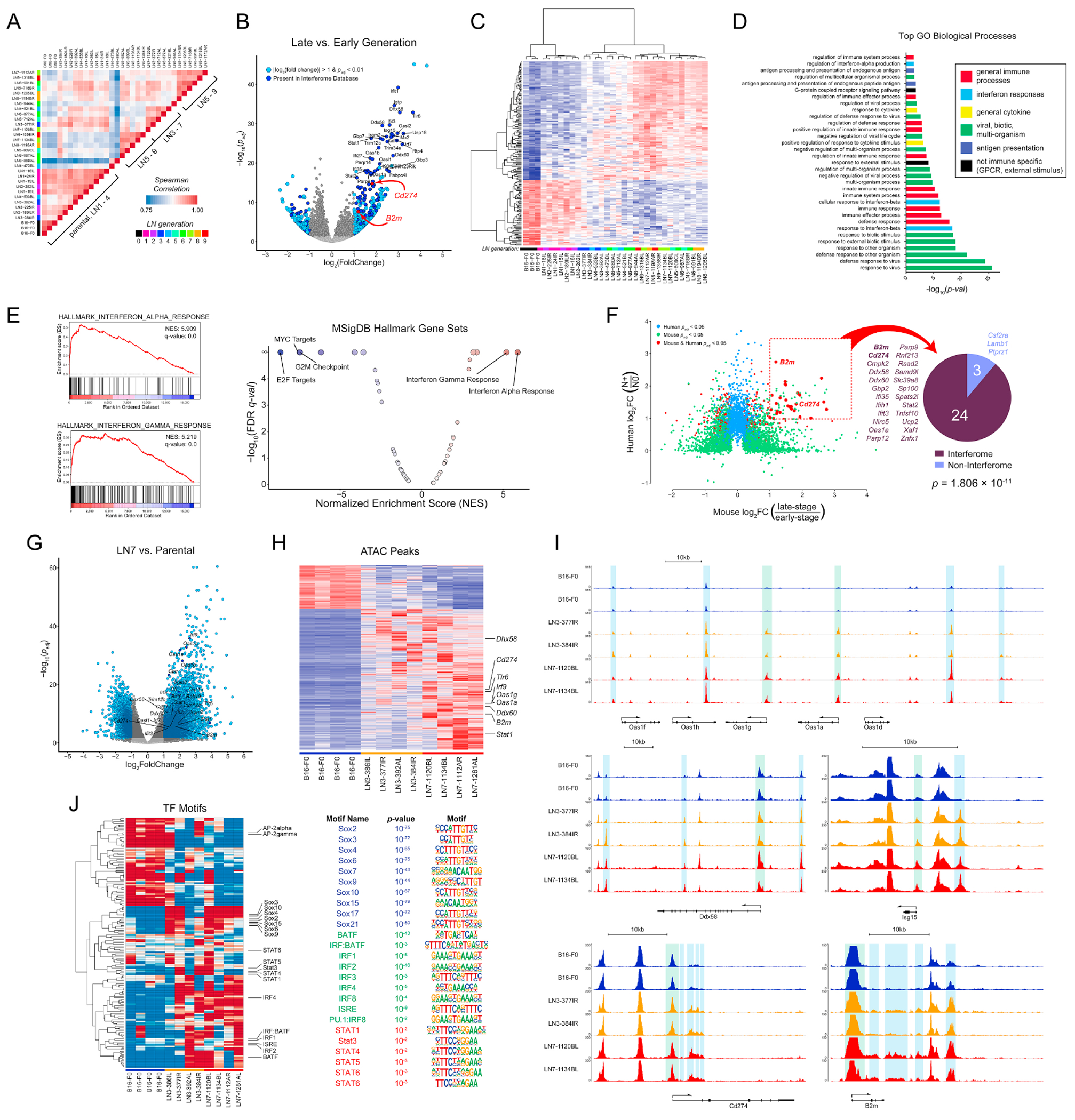
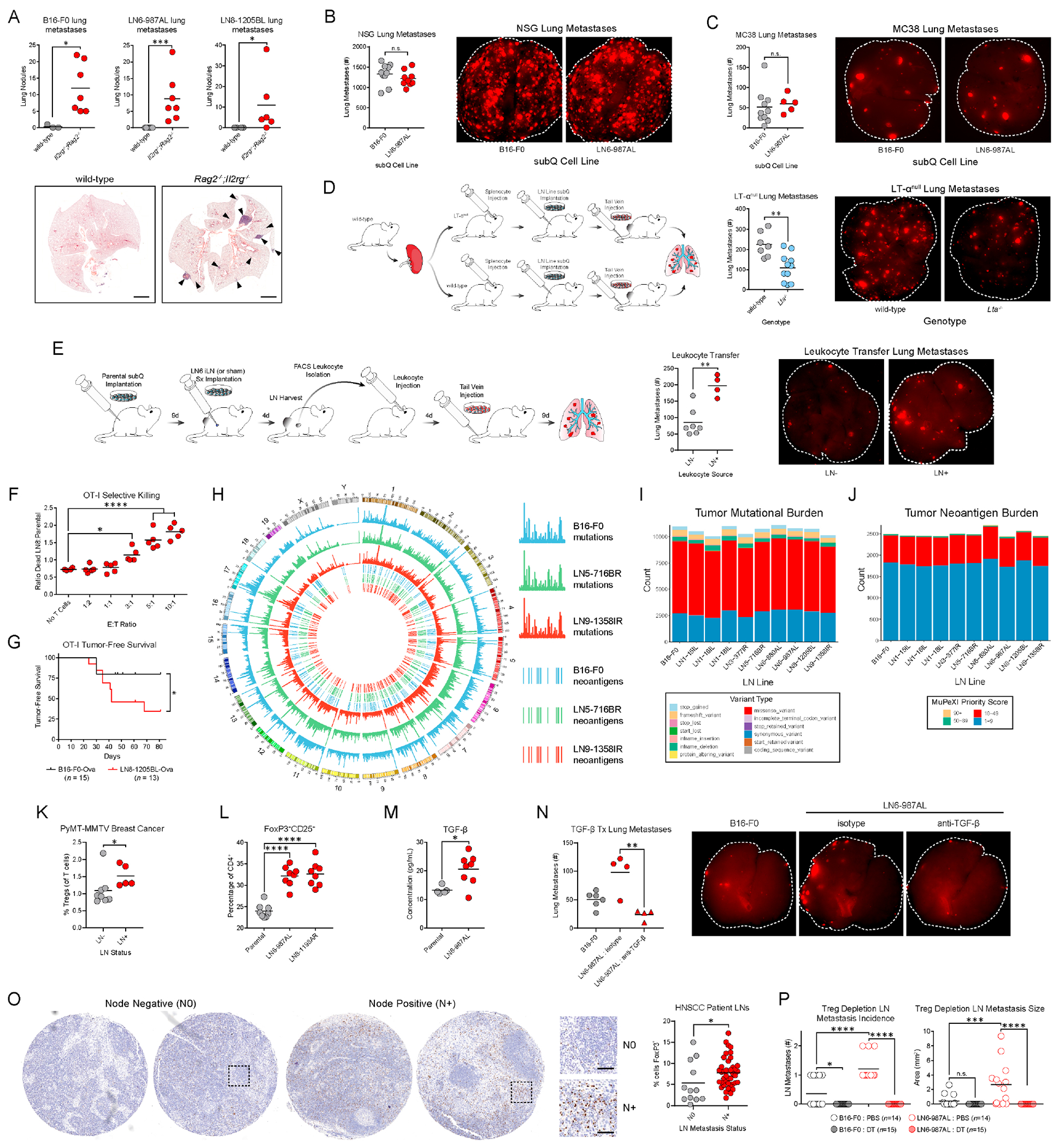
For many solid malignancies, lymph node (LN) involvement represents a harbinger of distant metastatic disease and, therefore, an important prognostic factor. Beyond its utility as a biomarker, whether and how LN metastasis plays an active role in shaping distant metastasis remains an open question. Here, we develop a syngeneic melanoma mouse model of LN metastasis to investigate how tumors spread to LNs and whether LN colonization influences metastasis to distant tissues. We show that an epigenetically instilled tumor-intrinsic interferon response program confers enhanced LN metastatic potential by enabling the evasion of NK cells and promoting LN colonization. LN metastases resist T cell-mediated cytotoxicity, induce antigen-specific regulatory T cells, and generate tumor-specific immune tolerance that subsequently facilitates distant tumor colonization. These effects extend to human cancers and other murine cancer models, implicating a conserved systemic mechanism by which malignancies spread to distant organs.
Keywords: ISGs; MHC-I; NK cells; PD-L1; Tregs; interferon; lymph nodes; metastasis; regulatory T cells; tolerance.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures





Comment in
-
Lymph node colonization and metastasis.Sci Signal. 2022 May 17;15(734):eadc9985. doi: 10.1126/scisignal.adc9985. Epub 2022 May 17. Sci Signal. 2022. PMID: 35580170
-
Lymph nodes tolerate tumours.Nat Rev Immunol. 2022 Jul;22(7):408-409. doi: 10.1038/s41577-022-00742-z. Nat Rev Immunol. 2022. PMID: 35610310 No abstract available.
-
Nodal interferon exposure drives tumour-specific tolerance.Nat Rev Cancer. 2022 Jul;22(7):377. doi: 10.1038/s41568-022-00489-8. Nat Rev Cancer. 2022. PMID: 35668152 No abstract available.
-
Lymph node metastasis fuels systemic disease.Trends Cancer. 2022 Aug;8(8):623-625. doi: 10.1016/j.trecan.2022.06.003. Epub 2022 Jun 15. Trends Cancer. 2022. PMID: 35717536
-
Node foretold: Cancer cells in lymph node rewire the immune system to enable further metastases.Cancer Cell. 2022 Aug 8;40(8):812-814. doi: 10.1016/j.ccell.2022.07.001. Epub 2022 Jul 14. Cancer Cell. 2022. PMID: 35839779
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
