

A novel small-molecule selective activator of homomeric GIRK4 channels
- PMID: 35525275
- PMCID: PMC9194863
- DOI: 10.1016/j.jbc.2022.102009
A novel small-molecule selective activator of homomeric GIRK4 channels
Abstract
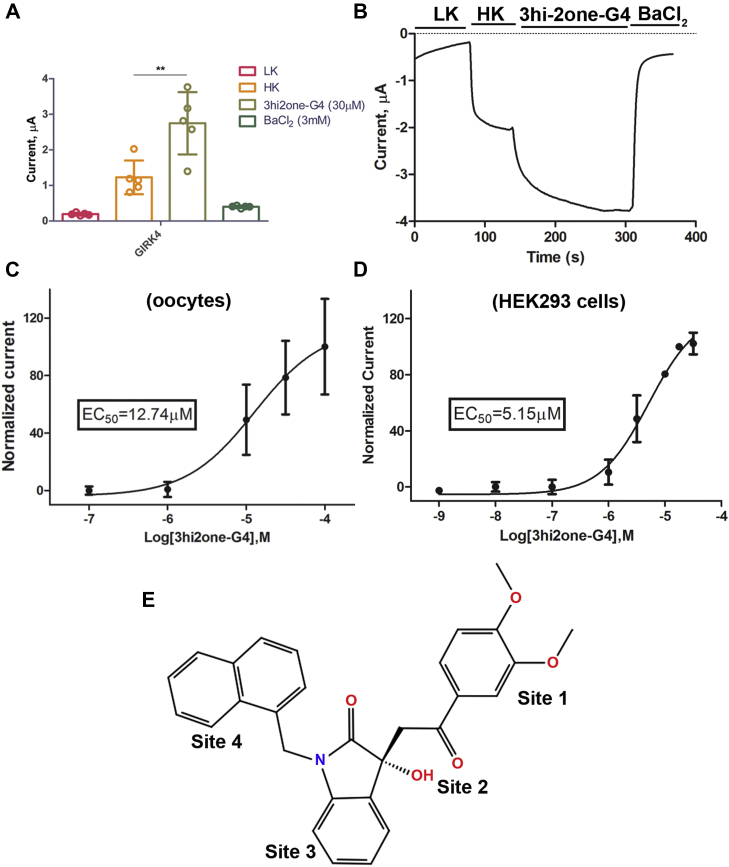
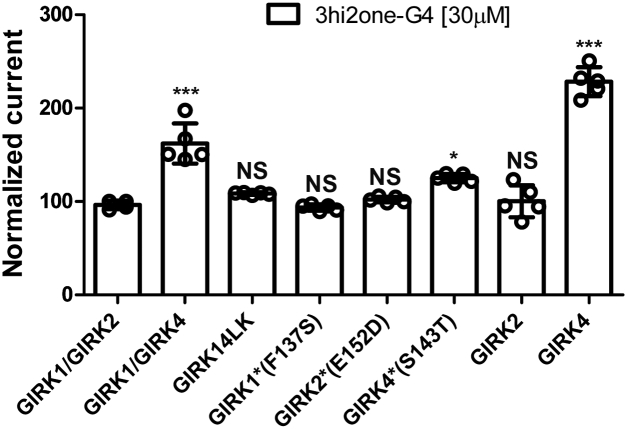
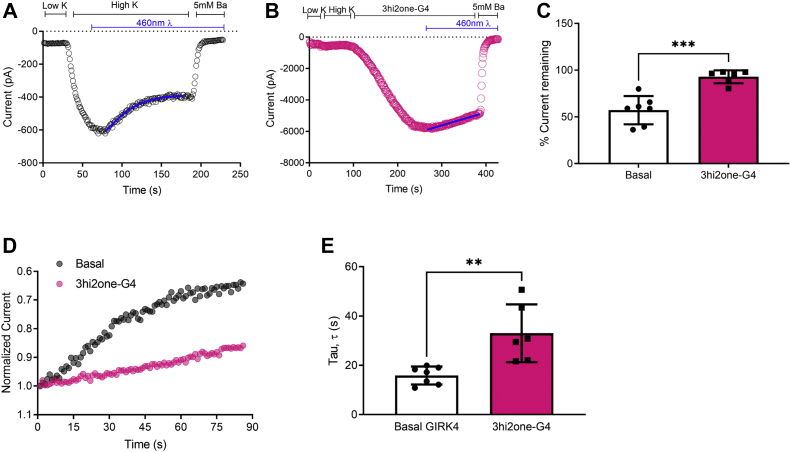
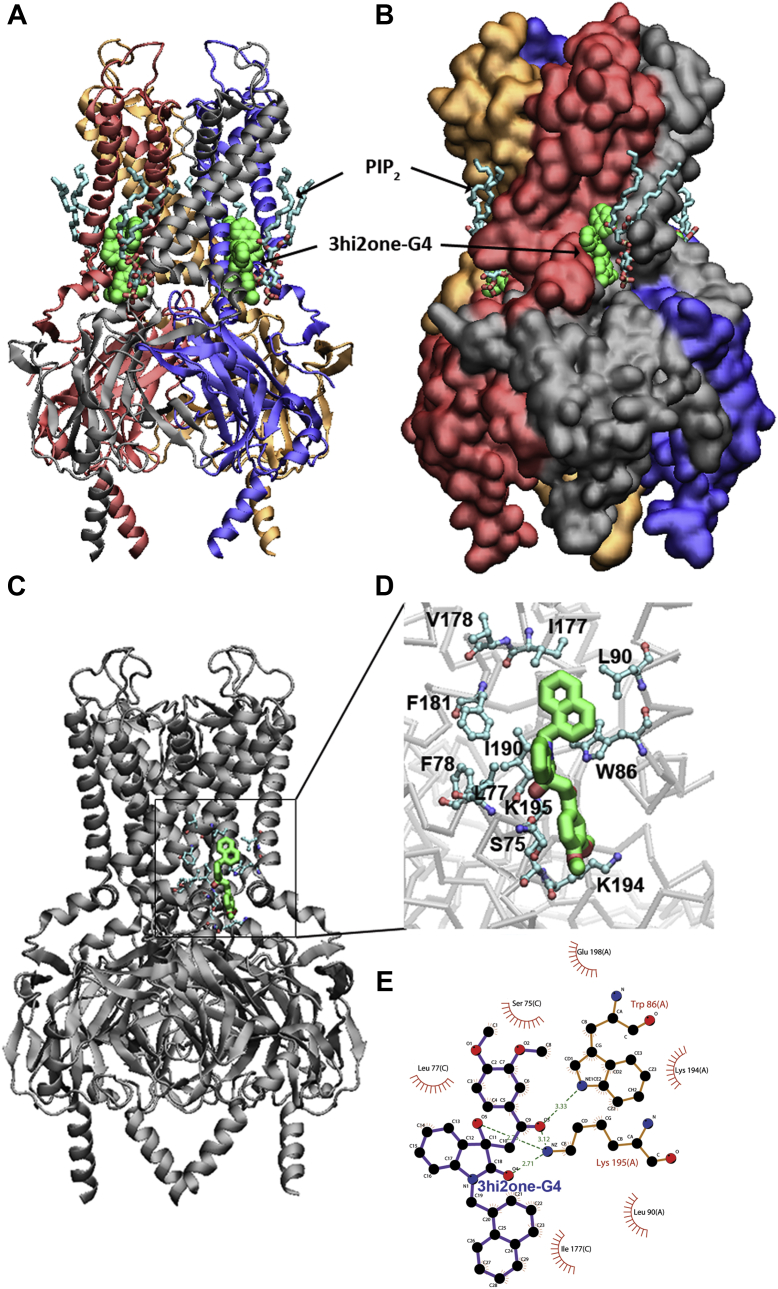
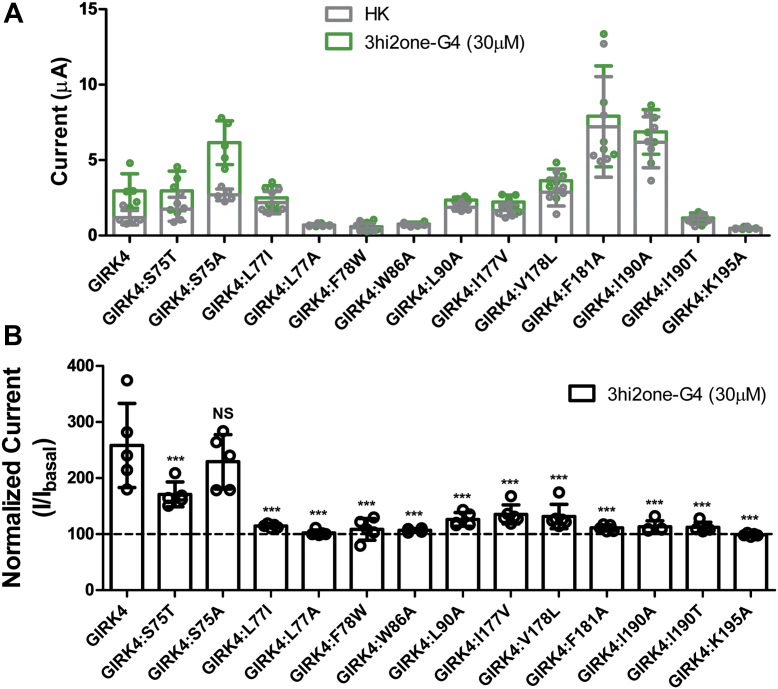
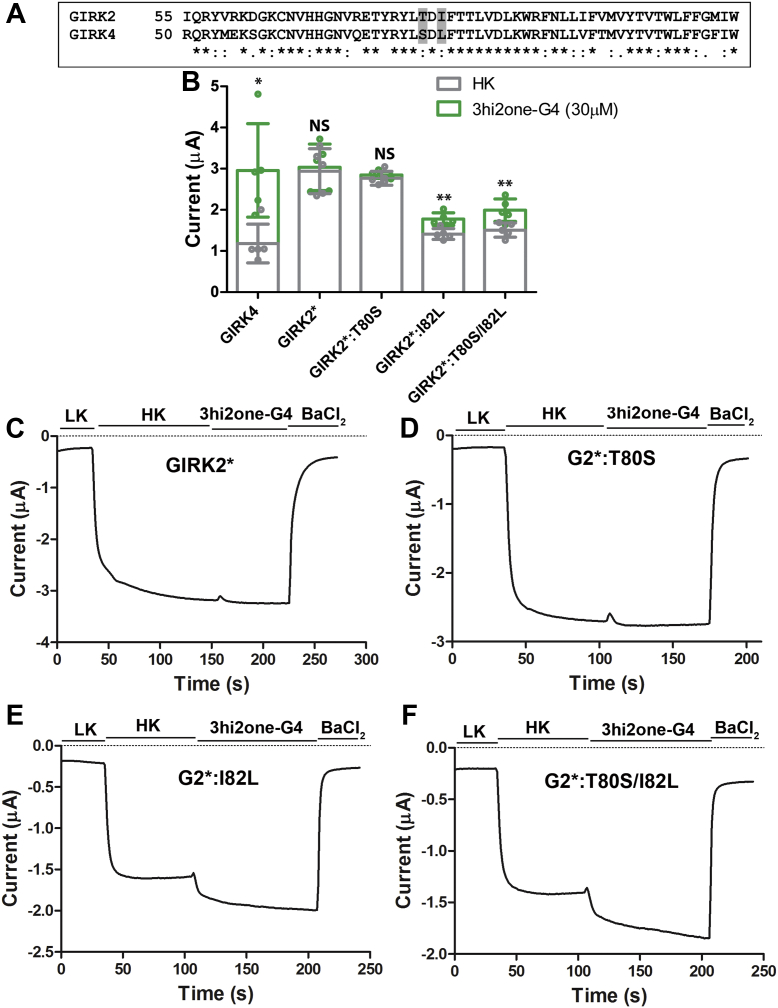
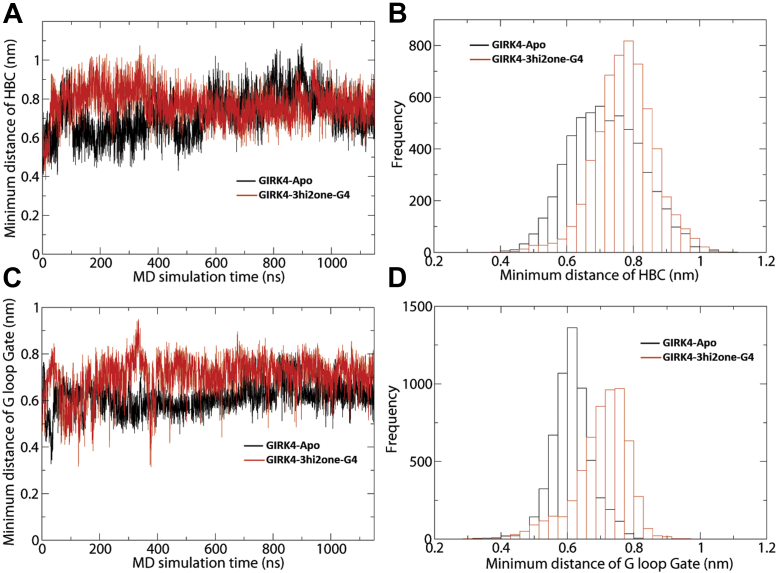
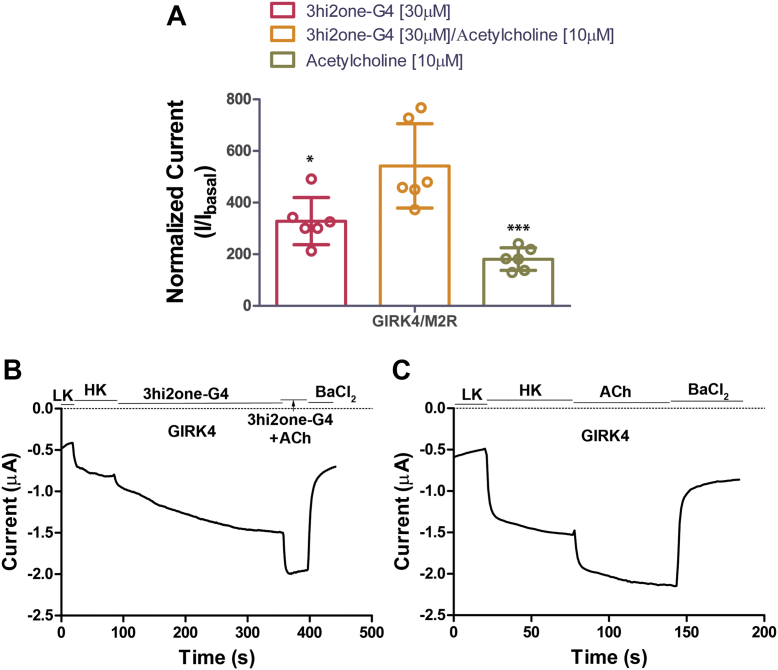
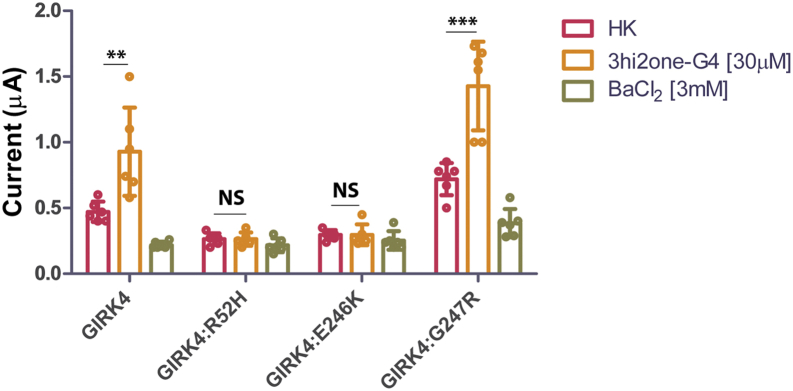
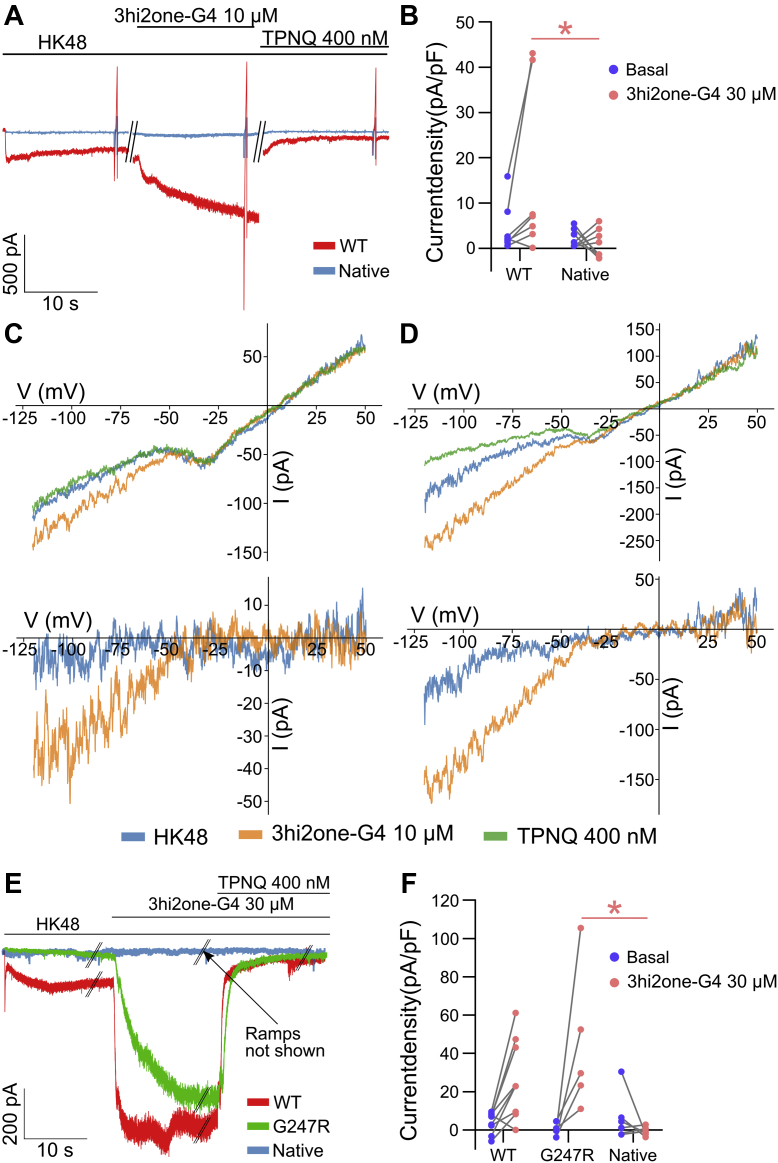
G protein-sensitive inwardly rectifying potassium (GIRK) channels are important pharmaceutical targets for neuronal, cardiac, and endocrine diseases. Although a number of GIRK channel modulators have been discovered in recent years, most lack selectivity. GIRK channels function as either homomeric (i.e., GIRK2 and GIRK4) or heteromeric (e.g., GIRK1/2, GIRK1/4, and GIRK2/3) tetramers. Activators, such as ML297, ivermectin, and GAT1508, have been shown to activate heteromeric GIRK1/2 channels better than GIRK1/4 channels with varying degrees of selectivity but not homomeric GIRK2 and GIRK4 channels. In addition, VU0529331 was discovered as the first homomeric GIRK channel activator, but it shows weak selectivity for GIRK2 over GIRK4 (or G4) homomeric channels. Here, we report the first highly selective small-molecule activator targeting GIRK4 homomeric channels, 3hi2one-G4 (3-[2-(3,4-dimethoxyphenyl)-2-oxoethyl]-3-hydroxy-1-(1-naphthylmethyl)-1,3-dihydro-2H-indol-2-one). We show that 3hi2one-G4 does not activate GIRK2, GIRK1/2, or GIRK1/4 channels. Using molecular modeling, mutagenesis, and electrophysiology, we analyzed the binding site of 3hi2one-G4 formed by the transmembrane 1, transmembrane 2, and slide helix regions of the GIRK4 channel, near the phosphatidylinositol-4,5-bisphosphate binding site, and show that it causes channel activation by strengthening channel-phosphatidylinositol-4,5-bisphosphate interactions. We also identify slide helix residue L77 in GIRK4, corresponding to residue I82 in GIRK2, as a major determinant of isoform-specific selectivity. We propose that 3hi2one-G4 could serve as a useful pharmaceutical probe in studying GIRK4 channel function and may also be pursued in drug optimization studies to tackle GIRK4-related diseases such as primary aldosteronism and late-onset obesity.
Keywords: Kir3 channels; drug–channel interaction; electrophysiology; molecular docking; molecular dynamics simulations; mutagenesis.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Cui M., Cantwell L., Zorn A., Logothetis D.E. Kir channel molecular physiology, pharmacology, and therapeutic implications. Handb. Exp. Pharmacol. 2021;267:277–356. - PubMed
-
- Wickman K., Nemec J., Gendler S.J., Clapham D.E. Abnormal heart rate regulation in GIRK4 knockout mice. Neuron. 1998;20:103–114. - PubMed
-
- Dobrev D., Friedrich A., Voigt N., Jost N., Wettwer E., Christ T., et al. The G protein-gated potassium current I(K,ACh) is constitutively active in patients with chronic atrial fibrillation. Circulation. 2005;112:3697–3706. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
