

Age-associated impairment of T cell immunity is linked to sex-dimorphic elevation of N-glycan branching
- PMID: 35528547
- PMCID: PMC9075523
- DOI: 10.1038/s43587-022-00187-y
Age-associated impairment of T cell immunity is linked to sex-dimorphic elevation of N-glycan branching
Abstract
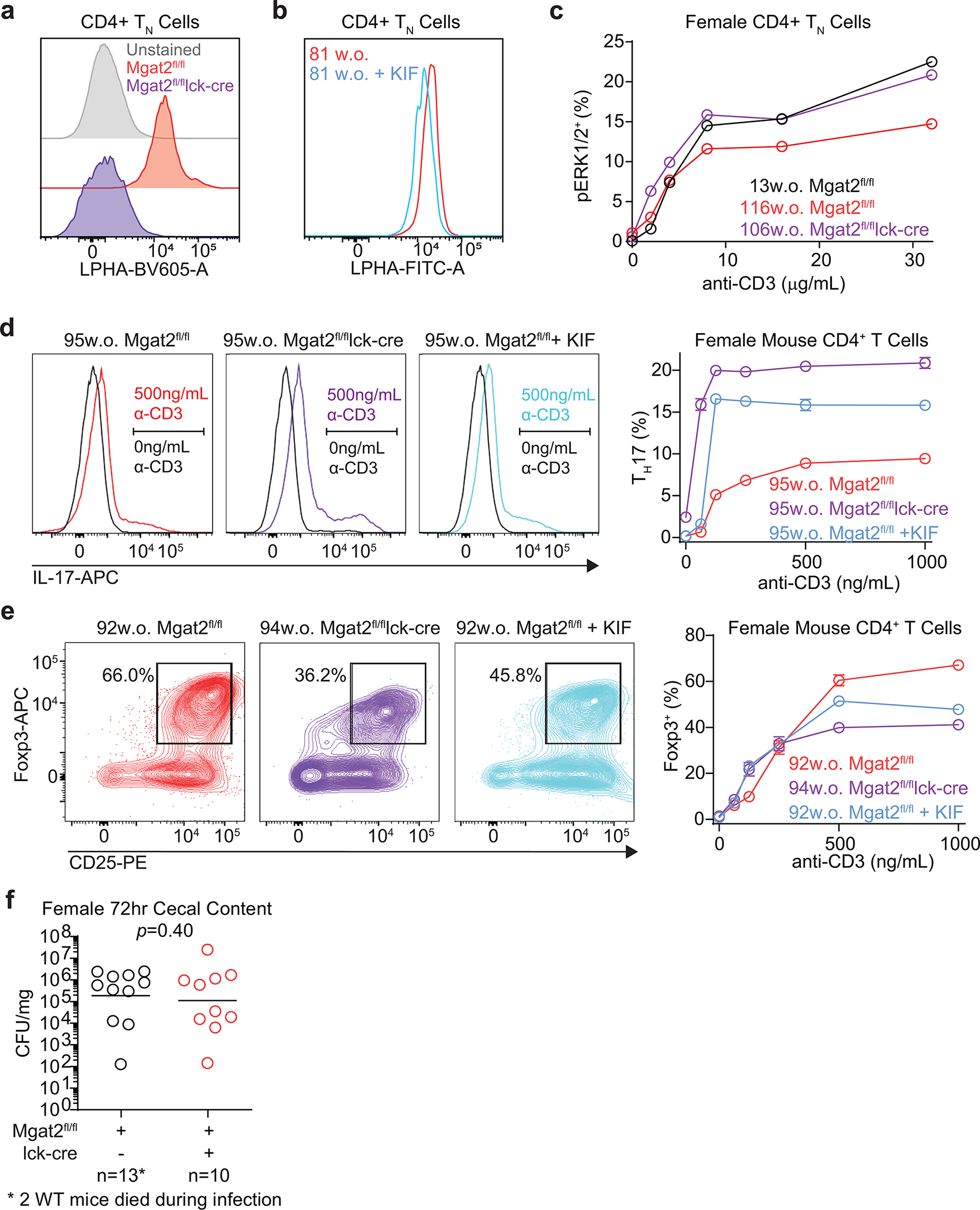
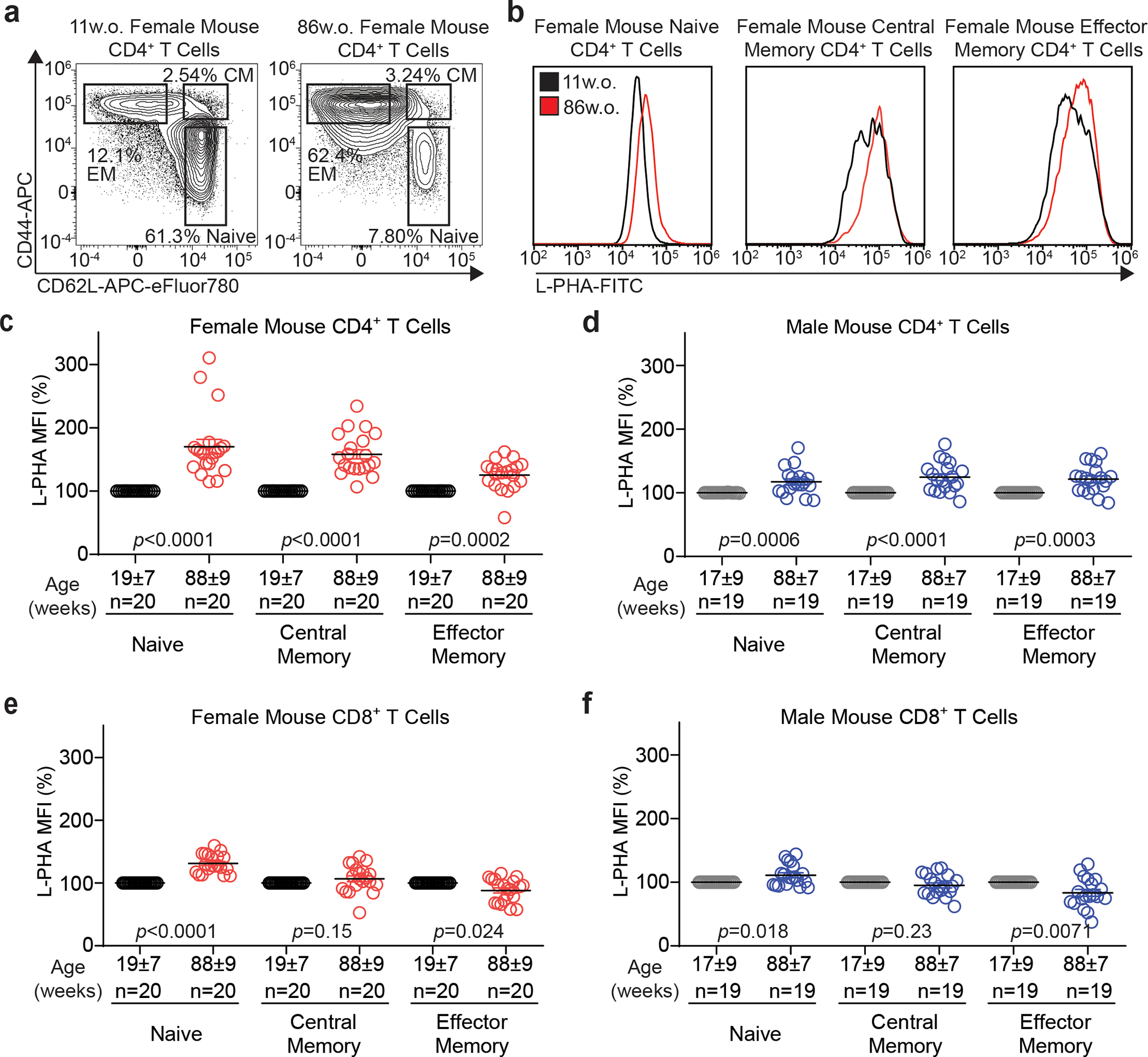
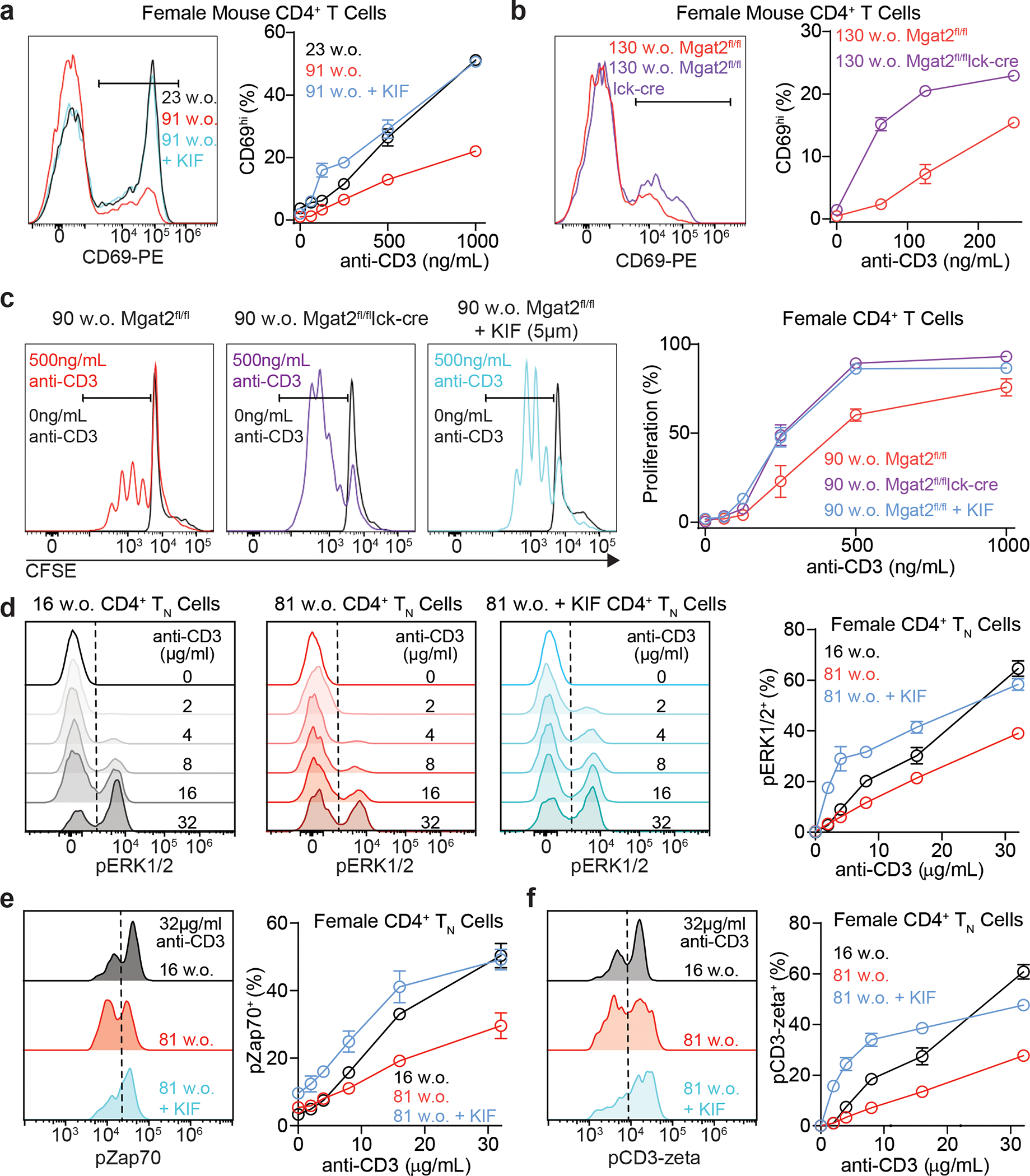
Impaired T cell immunity with aging increases mortality from infectious disease. The branching of Asparagine-linked glycans is a critical negative regulator of T cell immunity. Here we show that branching increases with age in females more than males, in naïve more than memory T cells, and in CD4+ more than CD8+ T cells. Female sex hormones and thymic output of naïve T cells (TN) decrease with age, however neither thymectomy nor ovariectomy altered branching. Interleukin-7 (IL-7) signaling was increased in old female more than male mouse TN cells, and triggered increased branching. N-acetylglucosamine, a rate-limiting metabolite for branching, increased with age in humans and synergized with IL-7 to raise branching. Reversing elevated branching rejuvenated T cell function and reduced severity of Salmonella infection in old female mice. These data suggest sex-dimorphic antagonistic pleiotropy, where IL-7 initially benefits immunity through TN maintenance but inhibits TN function by raising branching synergistically with age-dependent increases in N-acetylglucosamine.
Keywords: Immunosenescence; N-acetyglucosamine; N-glycan branching; N-glycosylation; T cell, infection; aging; immunity; interleukin-7.
Conflict of interest statement
Competing Interests Statement JD and MD are named as inventors on a patent application that describes GlcNAc as a biomarker for multiple sclerosis. JD and MD are named as inventors on a patent for use of GlcNAc in MS. The remaining authors declare no competing interests.
Figures









Comment in
-
Time isn't kind to female T-cells.Nat Aging. 2022 Mar;2(3):189-191. doi: 10.1038/s43587-022-00185-0. Epub 2022 Mar 18. Nat Aging. 2022. PMID: 36157102 Free PMC article.
References
-
- Centers for Disease, C. & Prevention. Estimates of deaths associated with seasonal influenza --- United States, 1976–2007. MMWR Morb Mortal Wkly Rep 59, 1057–1062 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
