

STING Is Required in Conventional Dendritic Cells for DNA Vaccine Induction of Type I T Helper Cell- Dependent Antibody Responses
- PMID: 35529875
- PMCID: PMC9072870
- DOI: 10.3389/fimmu.2022.861710
STING Is Required in Conventional Dendritic Cells for DNA Vaccine Induction of Type I T Helper Cell- Dependent Antibody Responses
Abstract
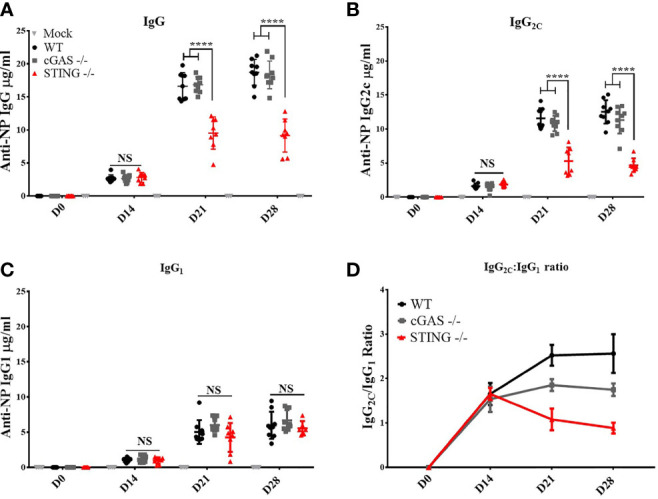
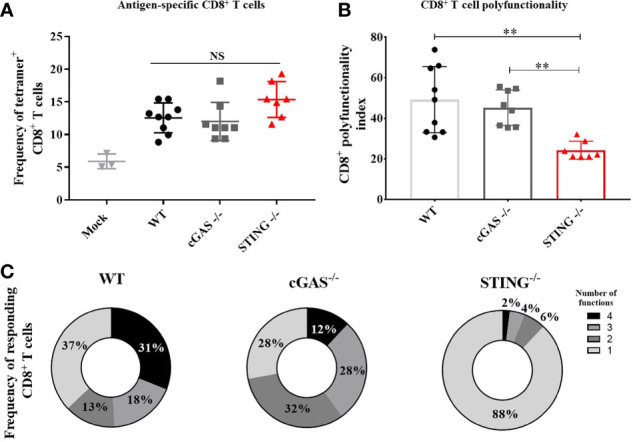
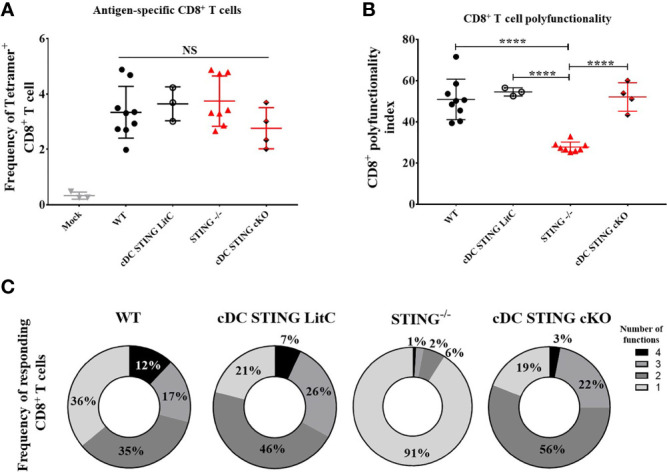
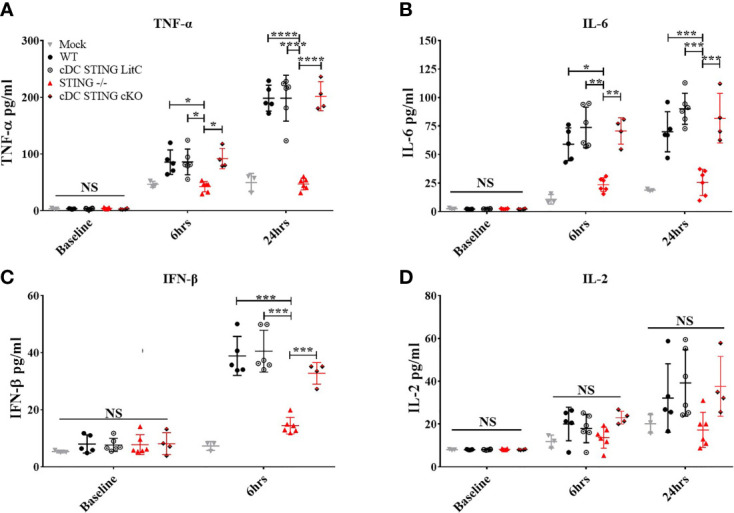
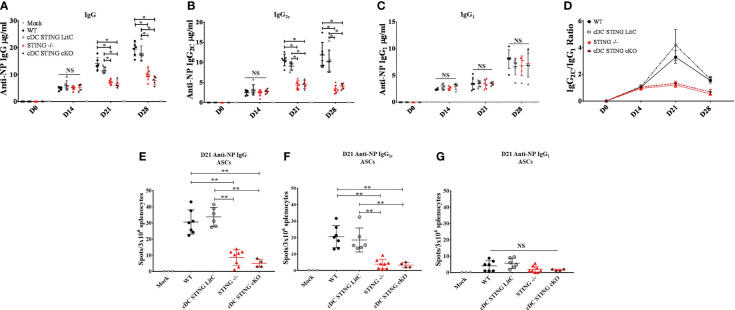
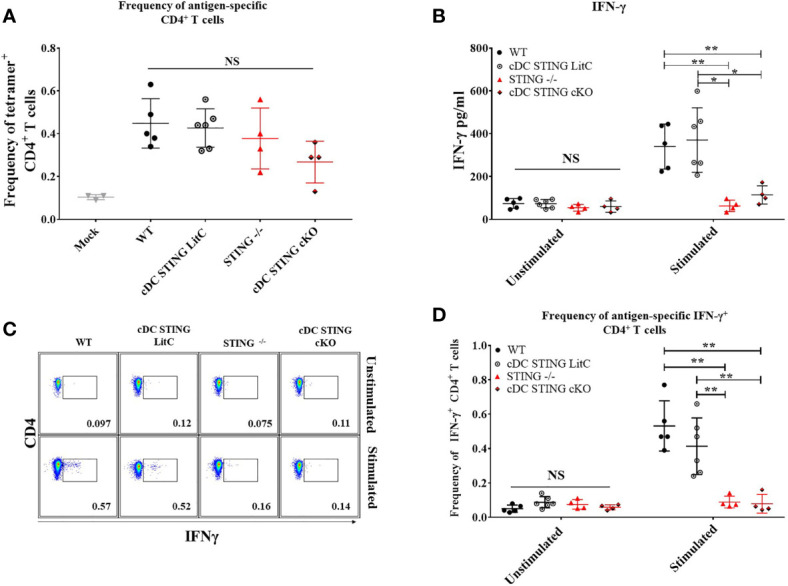
DNA vaccines elicit antibody, T helper cell responses and CD8+ T cell responses. Currently, little is known about the mechanism that DNA vaccines employ to induce adaptive immune responses. Prior studies have demonstrated that stimulator of interferon genes (STING) and conventional dendritic cells (cDCs) play critical roles in DNA vaccine induced antibody and T cell responses. STING activation by double stranded (dsDNA) sensing proteins initiate the production of type I interferon (IFN),but the DC-intrinsic effect of STING signaling is still unclear. Here, we investigated the role of STING within cDCs on DNA vaccine induction of antibody and T cell responses. STING knockout (STING-/- ) and conditional knockout mice that lack STING in cDCs (cDC STING cKO), were immunized intramuscularly with a DNA vaccine that expressed influenza A nucleoprotein (pNP). Both STING-/- and cDC STING cKO mice had significantly lower type I T helper (Th1) type antibody (anti-NP IgG2C) responses and lower frequencies of Th1 associated T cells (NP-specific IFN-γ+CD4+ T cells) post-immunization than wild type (WT) and cDC STING littermate control mice. In contrast, all mice had similar Th2-type NP-specific (IgG1) antibody titers. STING-/- mice developed significantly lower polyfunctional CD8+ T cells than WT, cDC STING cKO and cDC STING littermate control mice. These findings suggest that STING within cDCs mediates DNA vaccine induction of type I T helper responses including IFN-γ+CD4+ T cells, and Th1-type IgG2C antibody responses. The induction of CD8+ effector cell responses also require STING, but not within cDCs. These findings are the first to show that STING is required within cDCs to mediate DNA vaccine induced Th1 immune responses and provide new insight into the mechanism whereby DNA vaccines induce Th1 responses.
Keywords: DNA vaccine; STING; cGAS; dendritic cells; type I interferon.
Copyright © 2022 Ulrich-Lewis, Draves, Roe, O’Connor, Clark and Fuller.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
T follicular helper, but not Th1, cell differentiation in the absence of conventional dendritic cells.J Immunol. 2015 Jun 1;194(11):5187-99. doi: 10.4049/jimmunol.1401938. Epub 2015 Apr 27. J Immunol. 2015. PMID: 25917099
-
The Vaccine Adjuvant Chitosan Promotes Cellular Immunity via DNA Sensor cGAS-STING-Dependent Induction of Type I Interferons.Immunity. 2016 Mar 15;44(3):597-608. doi: 10.1016/j.immuni.2016.02.004. Epub 2016 Mar 2. Immunity. 2016. PMID: 26944200 Free PMC article.
-
Efficient Induction of Cytotoxic T Cells by Viral Vector Vaccination Requires STING-Dependent DC Functions.Front Immunol. 2020 Jul 16;11:1458. doi: 10.3389/fimmu.2020.01458. eCollection 2020. Front Immunol. 2020. PMID: 32765505 Free PMC article.
-
Mucosal immunity: regulation by helper T cells and a novel method for detection.J Biotechnol. 1996 Jan 26;44(1-3):209-16. doi: 10.1016/0168-1656(95)00095-X. J Biotechnol. 1996. PMID: 8717406 Review.
-
On taking the STING out of immune activation.J Leukoc Biol. 2018 Feb 12. doi: 10.1002/JLB.2MIR0917-383R. Online ahead of print. J Leukoc Biol. 2018. PMID: 29431896 Review.
Cited by
-
Poly I:C vaccination drives transient CXCL9 expression near B cell follicles in the lymph node through type-I and type-II interferon signaling.Cytokine. 2024 Nov;183:156731. doi: 10.1016/j.cyto.2024.156731. Epub 2024 Aug 20. Cytokine. 2024. PMID: 39168064
-
Multifaceted activation of STING axis upon Nipah and measles virus-induced syncytia formation.PLoS Pathog. 2024 Sep 16;20(9):e1012569. doi: 10.1371/journal.ppat.1012569. eCollection 2024 Sep. PLoS Pathog. 2024. PMID: 39283943 Free PMC article.
-
Harnessing innate immune pathways for therapeutic advancement in cancer.Signal Transduct Target Ther. 2024 Mar 25;9(1):68. doi: 10.1038/s41392-024-01765-9. Signal Transduct Target Ther. 2024. PMID: 38523155 Free PMC article. Review.
-
STING Protein-Based In Situ Vaccine Synergizes CD4+ T, CD8+ T, and NK Cells for Tumor Eradication.Adv Healthc Mater. 2023 Sep;12(24):e2300688. doi: 10.1002/adhm.202300688. Epub 2023 Apr 21. Adv Healthc Mater. 2023. PMID: 37015729 Free PMC article.
-
New insights for the development of efficient DNA vaccines.Microb Biotechnol. 2024 Nov;17(11):e70053. doi: 10.1111/1751-7915.70053. Microb Biotechnol. 2024. PMID: 39545748 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous