

Genetic and phylogenetic uncoupling of structure and function in human transmodal cortex
- PMID: 35534454
- PMCID: PMC9085871
- DOI: 10.1038/s41467-022-29886-1
Genetic and phylogenetic uncoupling of structure and function in human transmodal cortex
Abstract
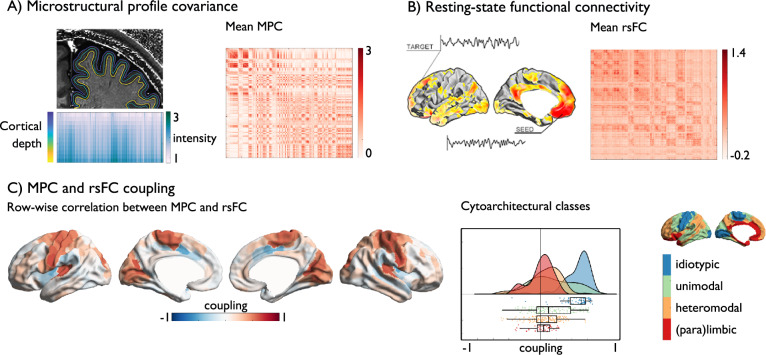
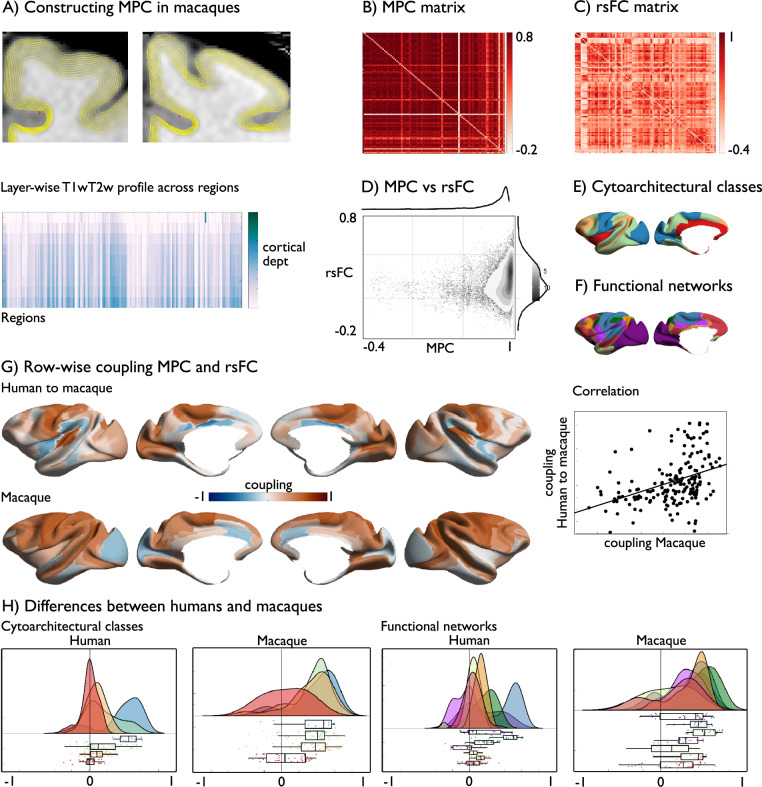
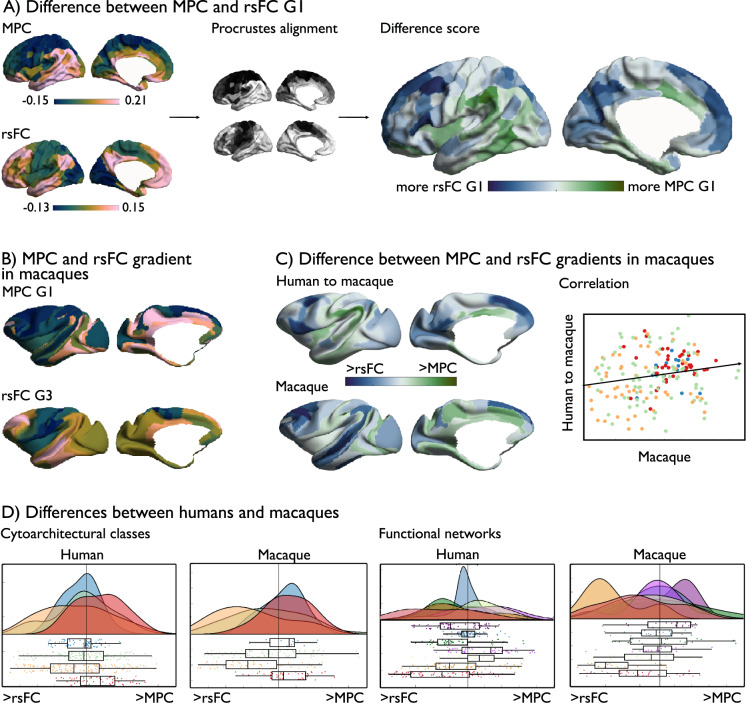
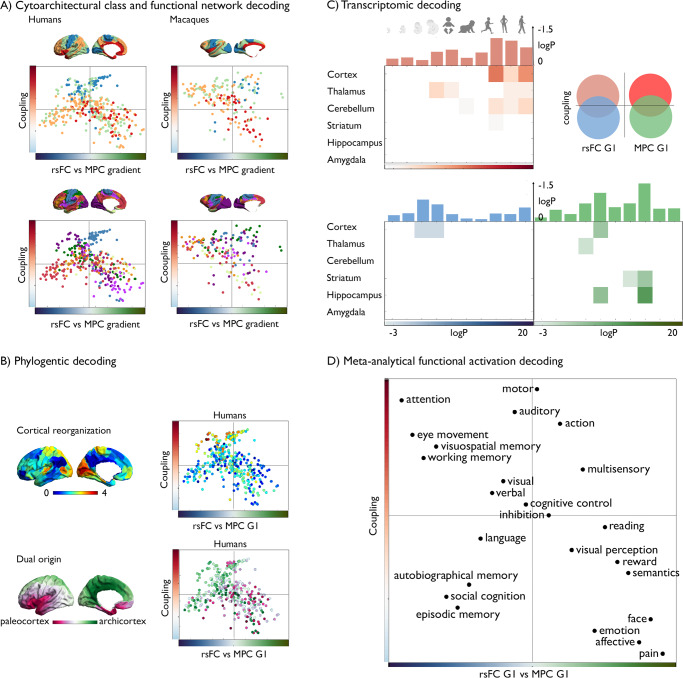
Brain structure scaffolds intrinsic function, supporting cognition and ultimately behavioral flexibility. However, it remains unclear how a static, genetically controlled architecture supports flexible cognition and behavior. Here, we synthesize genetic, phylogenetic and cognitive analyses to understand how the macroscale organization of structure-function coupling across the cortex can inform its role in cognition. In humans, structure-function coupling was highest in regions of unimodal cortex and lowest in transmodal cortex, a pattern that was mirrored by a reduced alignment with heritable connectivity profiles. Structure-function uncoupling in macaques had a similar spatial distribution, but we observed an increased coupling between structure and function in association cortices relative to humans. Meta-analysis suggested regions with the least genetic control (low heritable correspondence and different across primates) are linked to social-cognition and autobiographical memory. Our findings suggest that genetic and evolutionary uncoupling of structure and function in different transmodal systems may support the emergence of complex forms of cognition.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
A Comparative Perspective on the Cerebello-Cerebral System and Its Link to Cognition.Cerebellum. 2023 Dec;22(6):1293-1307. doi: 10.1007/s12311-022-01495-0. Epub 2022 Nov 23. Cerebellum. 2023. PMID: 36417091 Free PMC article. Review.
-
From sensation to cognition.Brain. 1998 Jun;121 ( Pt 6):1013-52. doi: 10.1093/brain/121.6.1013. Brain. 1998. PMID: 9648540 Review.
-
Modes of operation: A topographic neural gradient supporting stimulus dependent and independent cognition.Neuroimage. 2019 Feb 1;186:487-496. doi: 10.1016/j.neuroimage.2018.11.009. Epub 2018 Nov 14. Neuroimage. 2019. PMID: 30447291
-
Enhanced brain structure-function tethering in transmodal cortex revealed by high-frequency eigenmodes.Nat Commun. 2023 Oct 24;14(1):6744. doi: 10.1038/s41467-023-42053-4. Nat Commun. 2023. PMID: 37875493 Free PMC article.
-
Microstructural and functional gradients are increasingly dissociated in transmodal cortices.PLoS Biol. 2019 May 20;17(5):e3000284. doi: 10.1371/journal.pbio.3000284. eCollection 2019 May. PLoS Biol. 2019. PMID: 31107870 Free PMC article.
Cited by
-
A Comparative Perspective on the Cerebello-Cerebral System and Its Link to Cognition.Cerebellum. 2023 Dec;22(6):1293-1307. doi: 10.1007/s12311-022-01495-0. Epub 2022 Nov 23. Cerebellum. 2023. PMID: 36417091 Free PMC article. Review.
-
Diverging asymmetry of intrinsic functional organization in autism.Mol Psychiatry. 2023 Oct;28(10):4331-4341. doi: 10.1038/s41380-023-02220-x. Epub 2023 Aug 16. Mol Psychiatry. 2023. PMID: 37587246 Free PMC article.
-
Multimodal gradients unify local and global cortical organization.Nat Commun. 2025 Apr 25;16(1):3911. doi: 10.1038/s41467-025-59177-4. Nat Commun. 2025. PMID: 40280959 Free PMC article.
-
Group-common and individual-specific effects of structure-function coupling in human brain networks with graph neural networks.Imaging Neurosci (Camb). 2024 Dec 2;2:imag-2-00378. doi: 10.1162/imag_a_00378. eCollection 2024. Imaging Neurosci (Camb). 2024. PMID: 40800399 Free PMC article.
-
Personality traits vary in their association with brain activity across situations.Commun Biol. 2024 Nov 12;7(1):1498. doi: 10.1038/s42003-024-07061-0. Commun Biol. 2024. PMID: 39533085 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
