

Typical structure of rRNA coding genes in diplonemids points to two independent origins of the bizarre rDNA structures of euglenozoans
- PMID: 35534840
- PMCID: PMC9082867
- DOI: 10.1186/s12862-022-02014-9
Typical structure of rRNA coding genes in diplonemids points to two independent origins of the bizarre rDNA structures of euglenozoans
Abstract
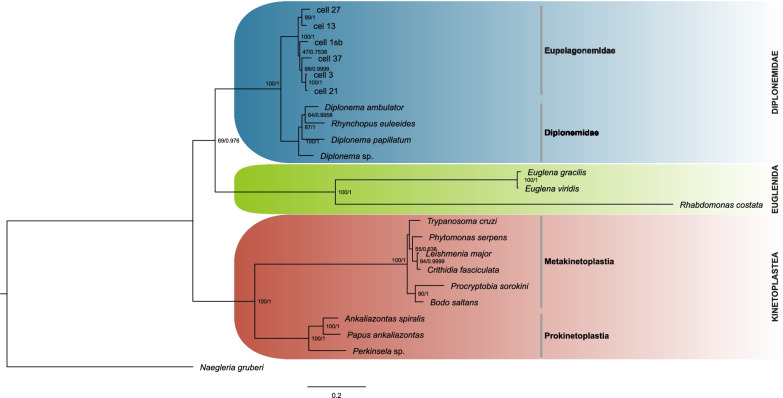
Background: Members of Euglenozoa (Discoba) are known for unorthodox rDNA organization. In Euglenida rDNA is located on extrachromosomal circular DNA. In Kinetoplastea and Euglenida the core of the large ribosomal subunit, typically formed by the 28S rRNA, consists of several smaller rRNAs. They are the result of the presence of additional internal transcribed spacers (ITSs) in the rDNA. Diplonemea is the third of the main groups of Euglenozoa and its members are known to be among the most abundant and diverse protists in the oceans. Despite that, the rRNA of only one diplonemid species, Diplonema papillatum, has been examined so far and found to exhibit continuous 28S rRNA. Currently, the rDNA organization has not been researched for any diplonemid. Herein we investigate the structure of rRNA genes in classical (Diplonemidae) and deep-sea diplonemids (Eupelagonemidae), representing the majority of known diplonemid diversity. The results fill the gap in knowledge about diplonemid rDNA and allow better understanding of the evolution of the fragmented structure of the rDNA in Euglenozoa.
Results: We used available genomic (culture and single-cell) sequencing data to assemble complete or almost complete rRNA operons for three classical and six deep-sea diplonemids. The rDNA sequences acquired for several euglenids and kinetoplastids were used to provide the background for the analysis. In all nine diplonemids, 28S rRNA seems to be contiguous, with no additional ITSs detected. Similarly, no additional ITSs were detected in basal prokinetoplastids. However, we identified five additional ITSs in the 28S rRNA of all analysed metakinetoplastids, and up to twelve in euglenids. Only three of these share positions, and they cannot be traced back to their common ancestor.
Conclusions: Presented results indicate that independent origin of additional ITSs in euglenids and kinetoplastids seems to be the most likely. The reason for such unmatched fragmentation remains unknown, but for some reason euglenozoan ribosomes appear to be prone to 28S rRNA fragmentation.
Keywords: Diplonemids; Euglenids; Euglenozoa; Internal transcribed spacer; Kinetoplastids; rDNA; rRNA; rRNA operon.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous