

Parallel Evolution of Ameloblastic scpp Genes in Bony and Cartilaginous Vertebrates
- PMID: 35535508
- PMCID: PMC9122587
- DOI: 10.1093/molbev/msac099
Parallel Evolution of Ameloblastic scpp Genes in Bony and Cartilaginous Vertebrates
Abstract
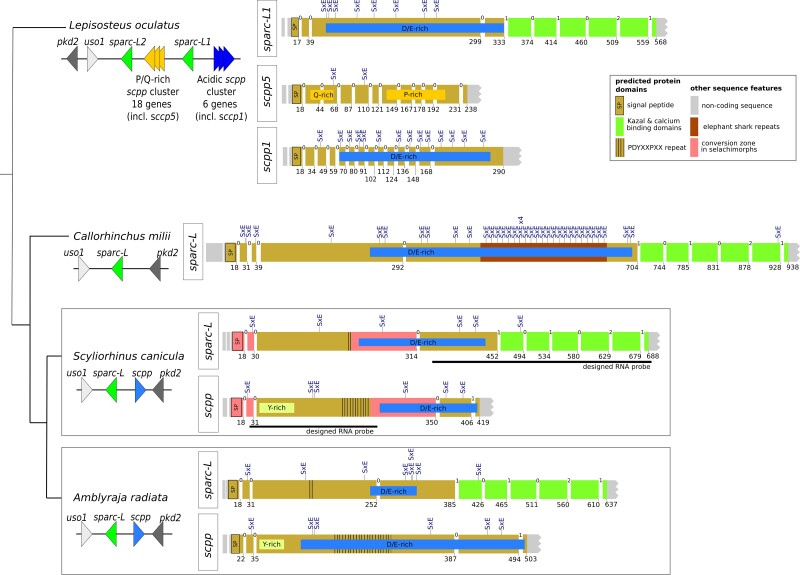
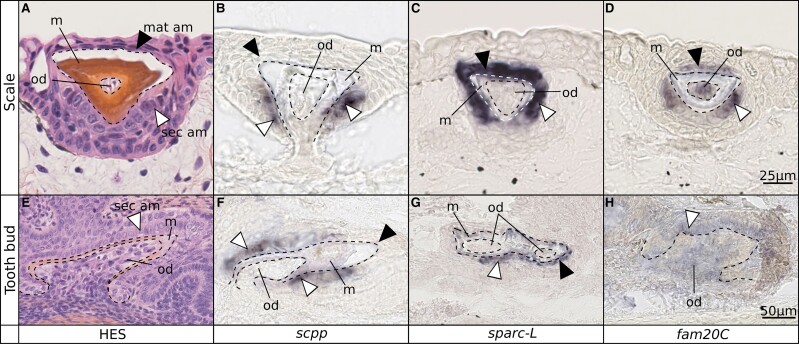
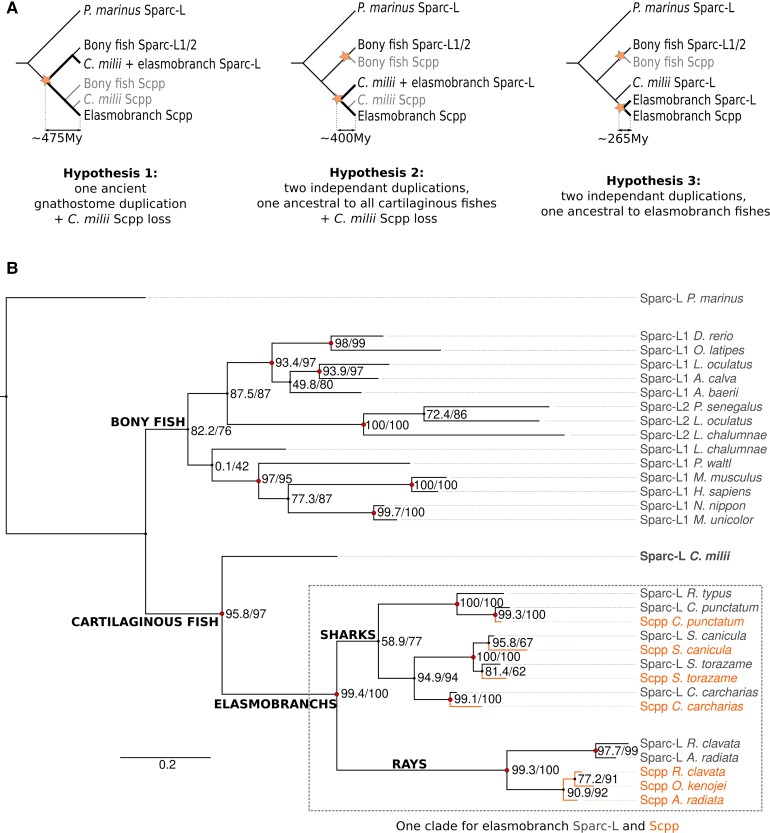
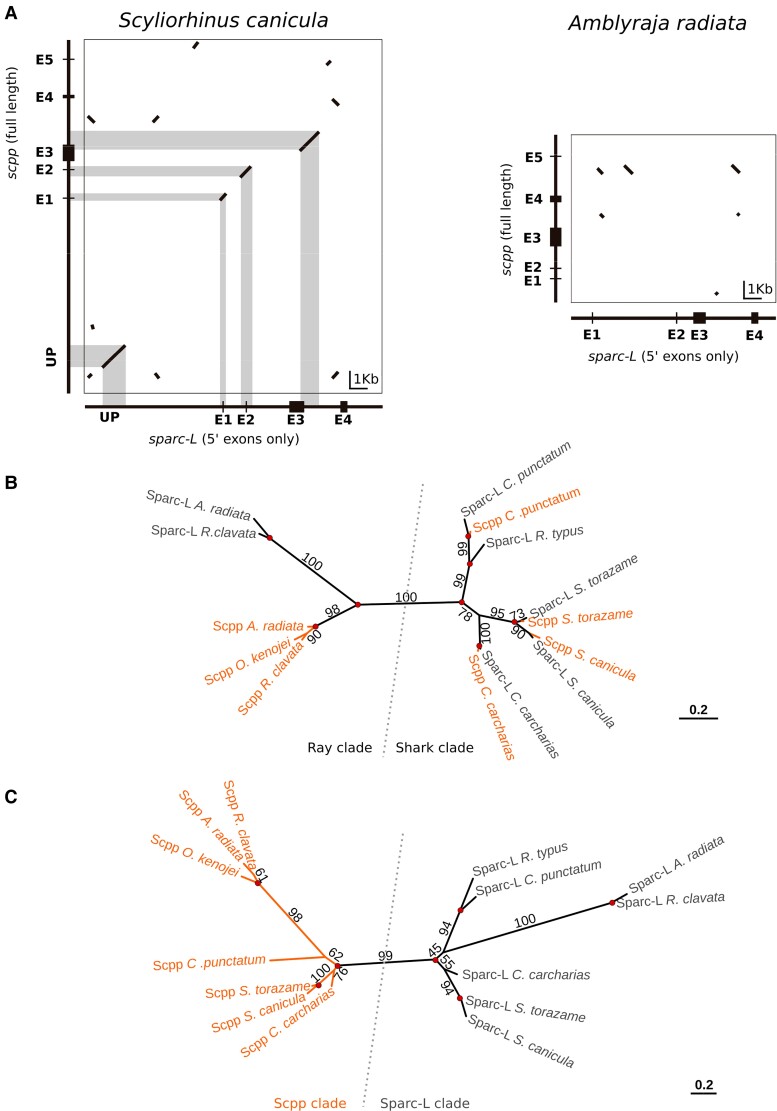
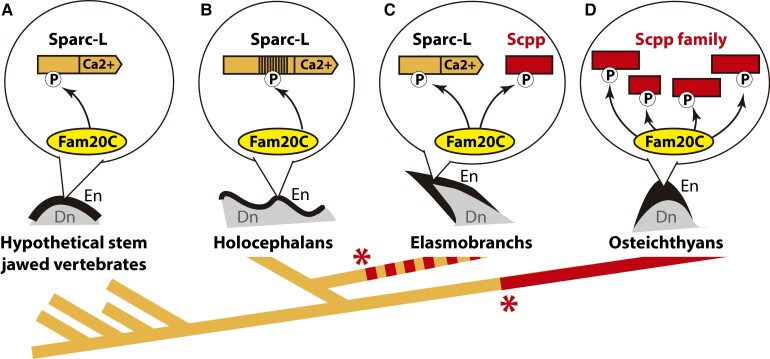
In bony vertebrates, skeletal mineralization relies on the secretory calcium-binding phosphoproteins (Scpp) family whose members are acidic extracellular proteins posttranslationally regulated by the Fam20°C kinase. As scpp genes are absent from the elephant shark genome, they are currently thought to be specific to bony fishes (osteichthyans). Here, we report a scpp gene present in elasmobranchs (sharks and rays) that evolved from local tandem duplication of sparc-L 5' exons and show that both genes experienced recent gene conversion in sharks. The elasmobranch scpp is remarkably similar to the osteichthyan scpp members as they share syntenic and gene structure features, code for a conserved signal peptide, tyrosine-rich and aspartate/glutamate-rich regions, and harbor putative Fam20°C phosphorylation sites. In addition, the catshark scpp is coexpressed with sparc-L and fam20°C in tooth and scale ameloblasts, similarly to some osteichthyan scpp genes. Despite these strong similarities, molecular clock and phylogenetic data demonstrate that the elasmobranch scpp gene originated independently from the osteichthyan scpp gene family. Our study reveals convergent events at the sparc-L locus in the two sister clades of jawed vertebrates, leading to parallel diversification of the skeletal biomineralization toolkit. The molecular evolution of sparc-L and its coexpression with fam20°C in catshark ameloblasts provides a unifying genetic basis that suggests that all convergent scpp duplicates inherited similar features from their sparc-L precursor. This conclusion supports a single origin for the hypermineralized outer odontode layer as produced by an ancestral developmental process performed by Sparc-L, implying the homology of the enamel and enameloid tissues in all vertebrates.
Keywords: Scyliorhinus canicula; fam20°C; scpp; sparc-L; ameloblasts; enamel; enameloid; gene conversion; genomic convergence; jawed vertebrate evolution.
© The Author(s) 2022. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures
References
-
- Berio F, Broyon M, Enault S NP, Lopez-Romero FA, Debiais-Thibaud M. 2021. Diversity and evolution of mineralized skeletal tissues in chondrichthyans. Front Ecol Evol. 9:1–19.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
