

α1B/D-adrenoceptors regulate chemokine receptor-mediated leukocyte migration via formation of heteromeric receptor complexes
- PMID: 35537053
- PMCID: PMC9171806
- DOI: 10.1073/pnas.2123511119
α1B/D-adrenoceptors regulate chemokine receptor-mediated leukocyte migration via formation of heteromeric receptor complexes
Abstract
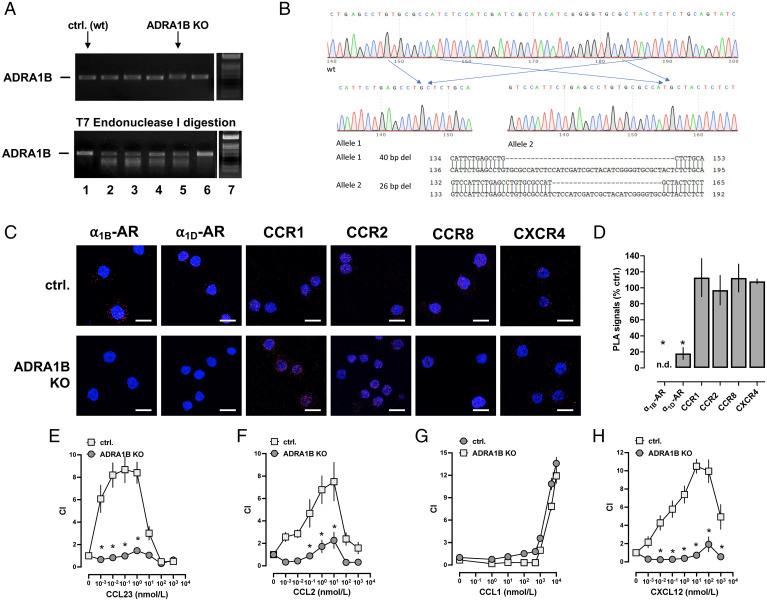
It is known that catecholamines regulate innate immune functions. The underlying mechanisms, however, are not well understood. Here we show that at least 20 members of the human chemokine receptor (CR) family heteromerize with one or more members of the α1-adrenergic receptor (AR) family in recombinant systems and that such heteromeric complexes are detectable in human monocytes and the monocytic leukemia cell line THP-1. Ligand binding to α1-ARs inhibited migration toward agonists of the CR heteromerization partners of α1B/D-ARs with high potency and 50 to 77% efficacy but did not affect migration induced by a noninteracting CR. Incomplete siRNA knockdown of α1B/D-ARs in THP-1 cells partially inhibited migration toward agonists of their CR heteromerization partners. Complete α1B-AR knockout via CRISPR-Cas9 gene editing in THP-1 cells (THP-1_ADRA1BKO) resulted in 82% reduction of α1D-AR expression and did not affect CR expression. Migration of THP-1_ADRA1BKO cells toward agonists of CR heteromerization partners of α1B/D-ARs was reduced by 82 to 95%. Our findings indicate that CR:α1B/D-AR heteromers are essential for normal function of CR heteromerization partners, provide a mechanism underlying neuroendocrine control of leukocyte trafficking, and offer opportunities to modulate leukocyte and/or cancer cell trafficking in disease processes.
Keywords: CRISPR-Cas9 gene editing; G protein–coupled receptor hetero-oligomers; chemokine receptors; chemotaxis; α1-adrenergic receptors.
Conflict of interest statement
The authors declare no competing interest.
Figures







Similar articles
-
α1-adrenoceptor ligands inhibit chemokine receptor heteromerization partners of α1B/D-adrenoceptors via interference with heteromer formation.Pharmacol Res. 2023 Apr;190:106730. doi: 10.1016/j.phrs.2023.106730. Epub 2023 Mar 15. Pharmacol Res. 2023. PMID: 36925091 Free PMC article.
-
α1-Adrenergic Receptors Function Within Hetero-Oligomeric Complexes With Atypical Chemokine Receptor 3 and Chemokine (C-X-C motif) Receptor 4 in Vascular Smooth Muscle Cells.J Am Heart Assoc. 2017 Aug 17;6(8):e006575. doi: 10.1161/JAHA.117.006575. J Am Heart Assoc. 2017. PMID: 28862946 Free PMC article.
-
Asymmetrical ligand-induced cross-regulation of chemokine (C-X-C motif) receptor 4 by α1-adrenergic receptors at the heteromeric receptor complex.Sci Rep. 2018 Feb 9;8(1):2730. doi: 10.1038/s41598-018-21096-4. Sci Rep. 2018. PMID: 29426850 Free PMC article.
-
Cardiac and Vascular α1-Adrenoceptors in Congestive Heart Failure: A Systematic Review.Cells. 2020 Nov 4;9(11):2412. doi: 10.3390/cells9112412. Cells. 2020. PMID: 33158106 Free PMC article.
-
Recent progress in alpha1-adrenergic receptor research.Acta Pharmacol Sin. 2005 Nov;26(11):1281-7. doi: 10.1111/j.1745-7254.2005.00224.x. Acta Pharmacol Sin. 2005. PMID: 16225747 Review.
Cited by
-
α1-adrenoceptor ligands inhibit chemokine receptor heteromerization partners of α1B/D-adrenoceptors via interference with heteromer formation.Pharmacol Res. 2023 Apr;190:106730. doi: 10.1016/j.phrs.2023.106730. Epub 2023 Mar 15. Pharmacol Res. 2023. PMID: 36925091 Free PMC article.
-
Sympathetic System in Wound Healing: Multistage Control in Normal and Diabetic Skin.Int J Mol Sci. 2023 Jan 20;24(3):2045. doi: 10.3390/ijms24032045. Int J Mol Sci. 2023. PMID: 36768369 Free PMC article. Review.
-
α1-Adrenergic Receptors: Insights into Potential Therapeutic Opportunities for COVID-19, Heart Failure, and Alzheimer's Disease.Int J Mol Sci. 2023 Feb 20;24(4):4188. doi: 10.3390/ijms24044188. Int J Mol Sci. 2023. PMID: 36835598 Free PMC article. Review.
-
Chemokine receptor hetero-oligomers regulate monocyte chemotaxis.Life Sci Alliance. 2024 May 23;7(8):e202402657. doi: 10.26508/lsa.202402657. Print 2024 Aug. Life Sci Alliance. 2024. PMID: 38782603 Free PMC article.
-
Recent advances in the role of neuroregulation in skin wound healing.Burns Trauma. 2025 Jan 27;13:tkae072. doi: 10.1093/burnst/tkae072. eCollection 2025. Burns Trauma. 2025. PMID: 39872039 Free PMC article. Review.
References
-
- Dhabhar F. S., Effects of stress on immune function: The good, the bad, and the beautiful. Immunol. Res. 58, 193–210 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
