

Synaptotagmin-1 is a bidirectional Ca2+ sensor for neuronal endocytosis
- PMID: 35537054
- PMCID: PMC9171800
- DOI: 10.1073/pnas.2111051119
Synaptotagmin-1 is a bidirectional Ca2+ sensor for neuronal endocytosis
Abstract
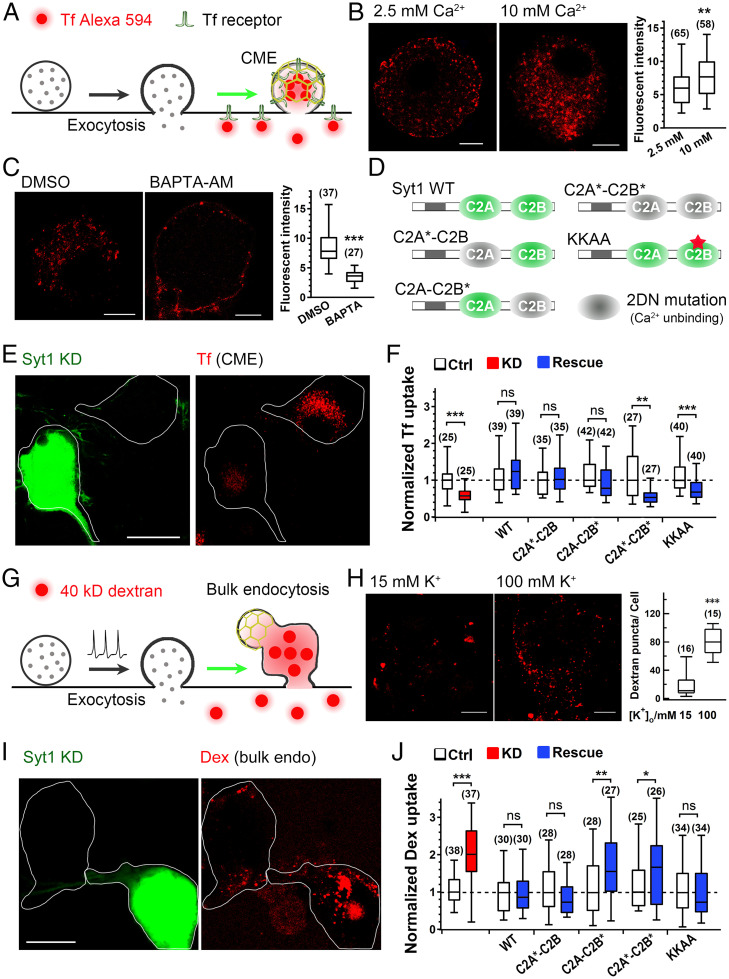
Exocytosis and endocytosis are tightly coupled. In addition to initiating exocytosis, Ca2+ plays critical roles in exocytosis–endocytosis coupling in neurons and nonneuronal cells. Both positive and negative roles of Ca2+ in endocytosis have been reported; however, Ca2+ inhibition in endocytosis remains debatable with unknown mechanisms. Here, we show that synaptotagmin-1 (Syt1), the primary Ca2+ sensor initiating exocytosis, plays bidirectional and opposite roles in exocytosis–endocytosis coupling by promoting slow, small-sized clathrin-mediated endocytosis but inhibiting fast, large-sized bulk endocytosis. Ca2+-binding ability is required for Syt1 to regulate both types of endocytic pathways, the disruption of which leads to inefficient vesicle recycling under mild stimulation and excessive membrane retrieval following intense stimulation. Ca2+-dependent membrane tubulation may explain the opposite endocytic roles of Syt1 and provides a general membrane-remodeling working model for endocytosis determination. Thus, Syt1 is a primary bidirectional Ca2+ sensor facilitating clathrin-mediated endocytosis but clamping bulk endocytosis, probably by manipulating membrane curvature to ensure both efficient and precise coupling of endocytosis to exocytosis.
Keywords: Ca2+; bulk endocytosis; clathrin-mediated endocytosis; membrane tubulation; synaptotagmin.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
-
- Leitz J., Kavalali E. T., Ca2+ dependence of synaptic vesicle endocytosis. Neuroscientist 22, 464–476 (2016). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
