

Melanoma cells with diverse invasive potential differentially induce the activation of normal human fibroblasts
- PMID: 35538545
- PMCID: PMC9092709
- DOI: 10.1186/s12964-022-00871-x
Melanoma cells with diverse invasive potential differentially induce the activation of normal human fibroblasts
Abstract
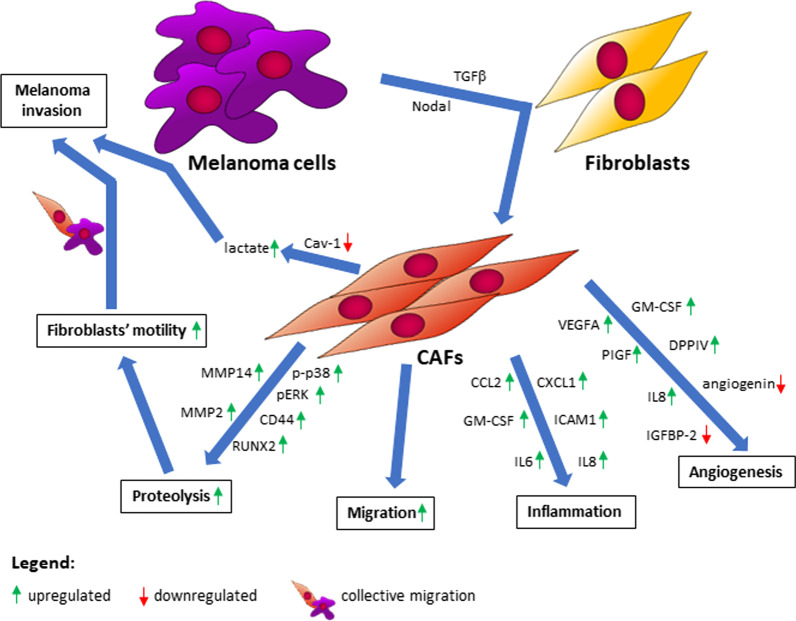
Background: The tumor microenvironment consists of stromal cells, extracellular matrix, and physicochemical properties (e.g., oxygenation, acidification). An important element of the tumor niche are cancer-associated fibroblasts (CAFs). They may constitute up to 80% of the tumor mass and share some features with myofibroblasts involved in the process of wound healing. CAFs can facilitate cancer progression. However, their interaction with melanoma cells is still poorly understood.
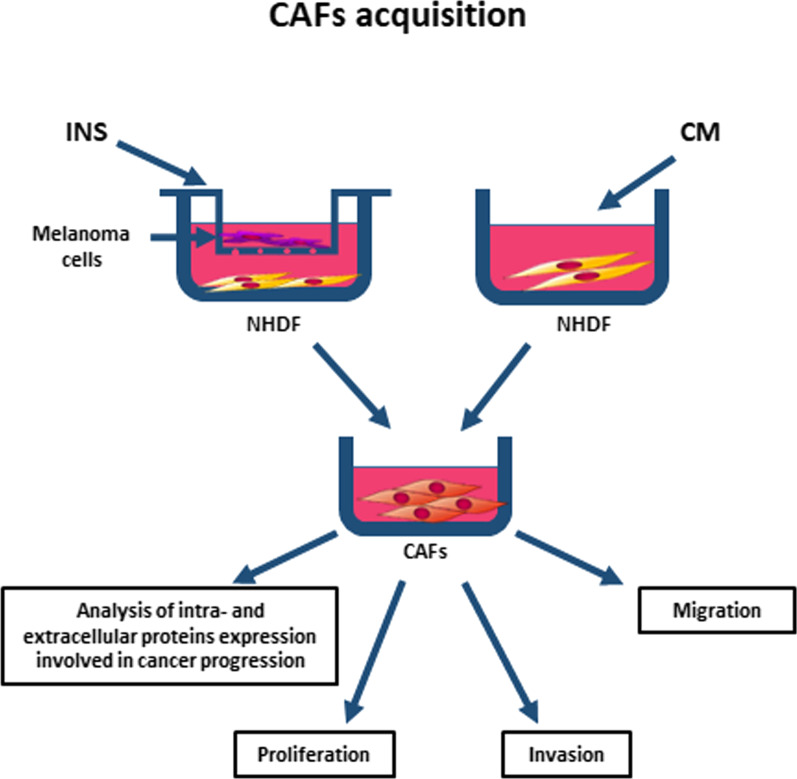
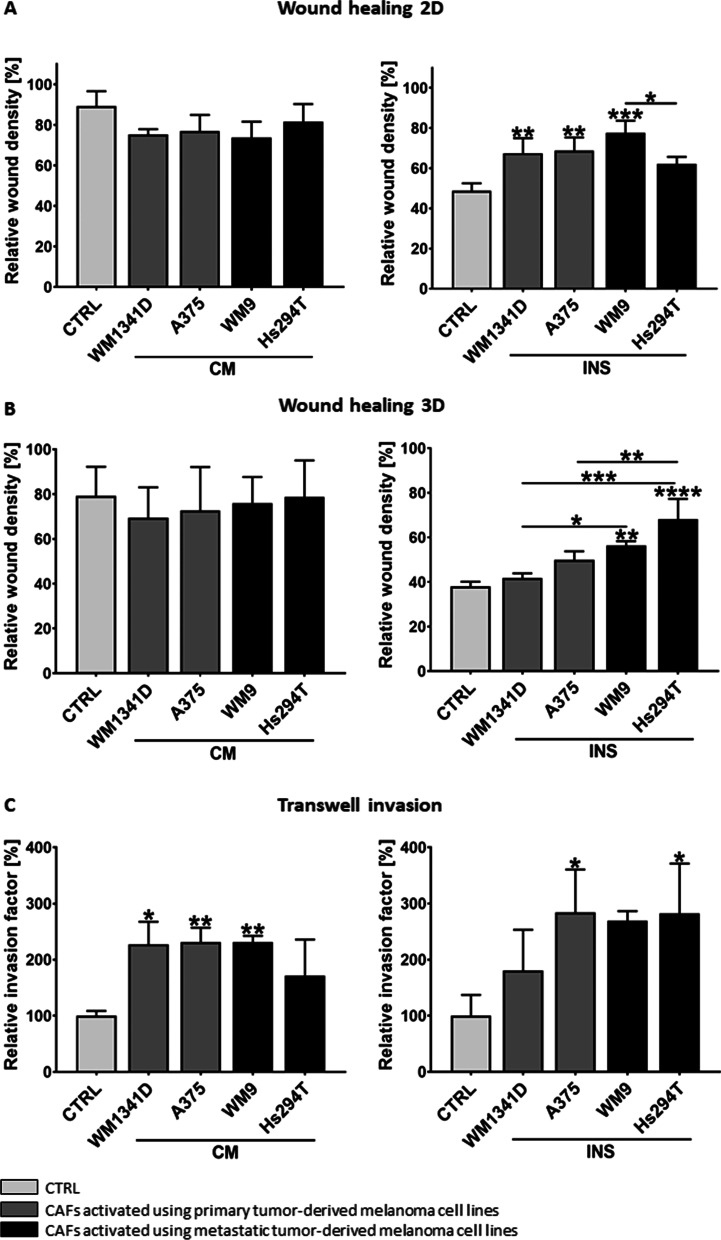
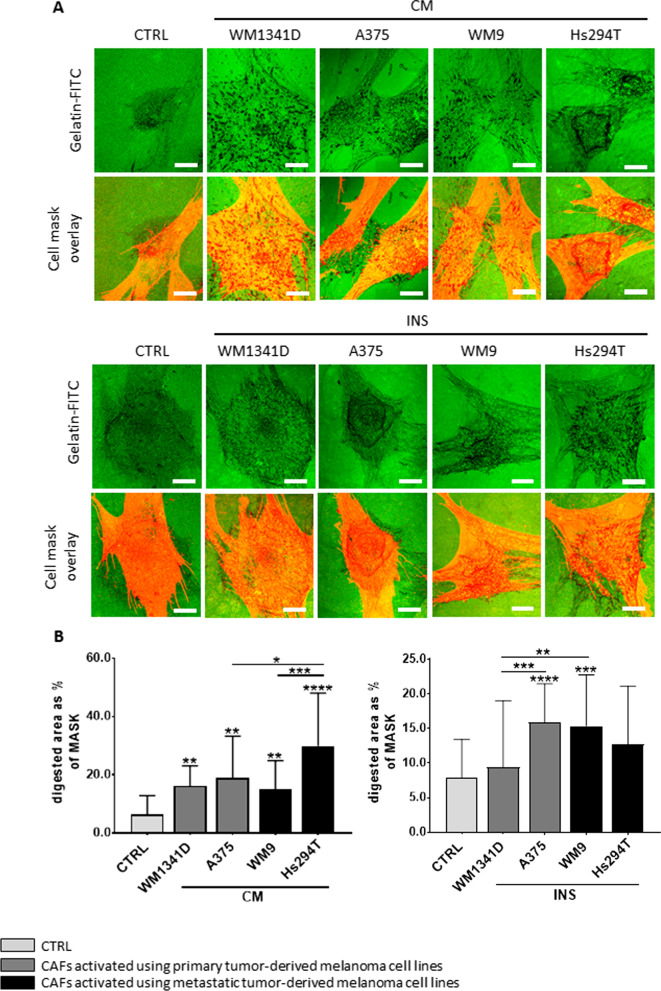
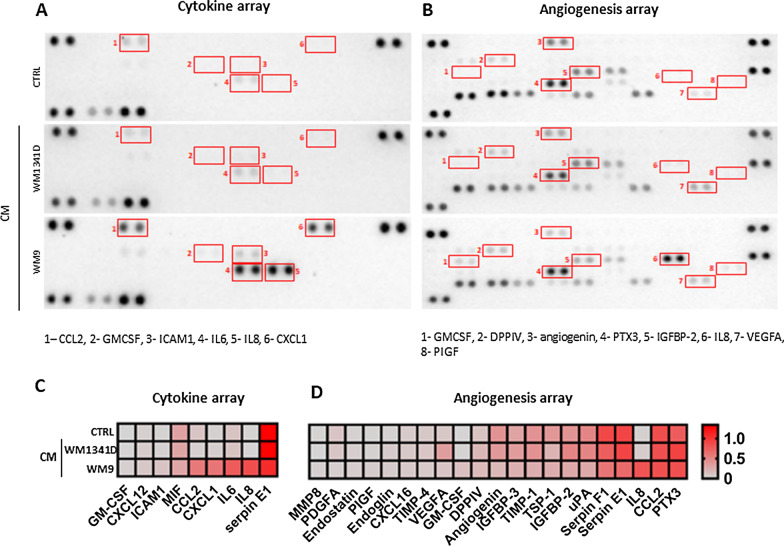
Methods: We obtained CAFs using conditioned media derived from primary and metastatic melanoma cells, and via co-culture with melanoma cells on Transwell inserts. Using 2D and 3D wound healing assays and Transwell invasion method we evaluated CAFs' motile activities, while coverslips with FITC-labeled gelatin, gelatin zymography, and fluorescence-based activity assay were employed to determine the proteolytic activity of the examined cells. Western Blotting method was used for the identification of CAFs' markers as well as estimation of the mediators of MMPs' (matrix metalloproteinases) expression levels. Lastly, CAFs' secretome was evaluated with cytokine and angiogenesis proteomic arrays, and lactate chemiluminescence-based assay.
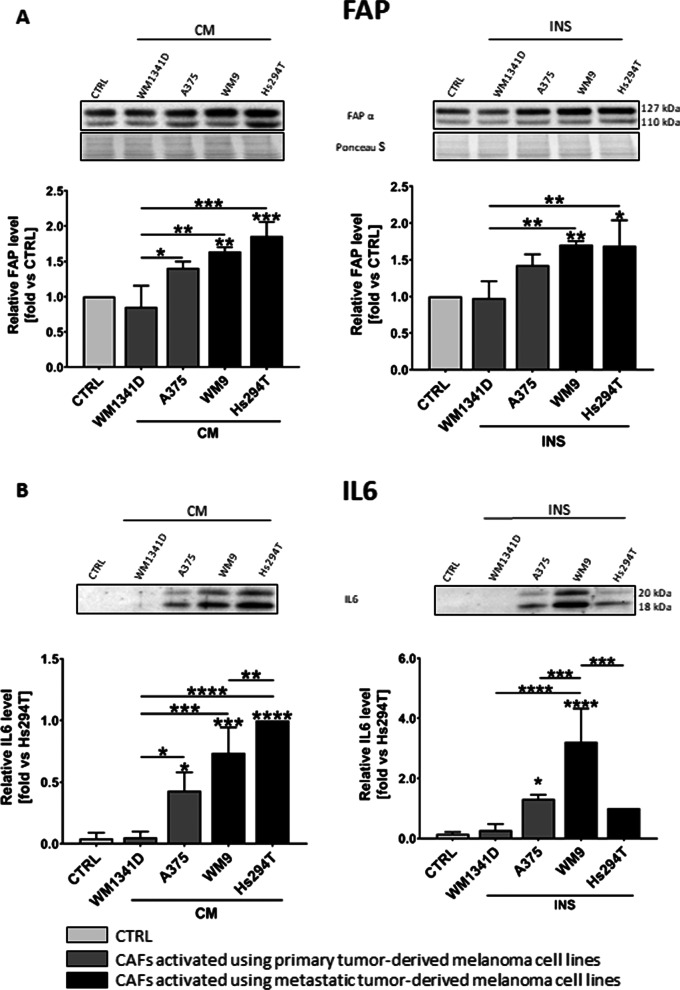
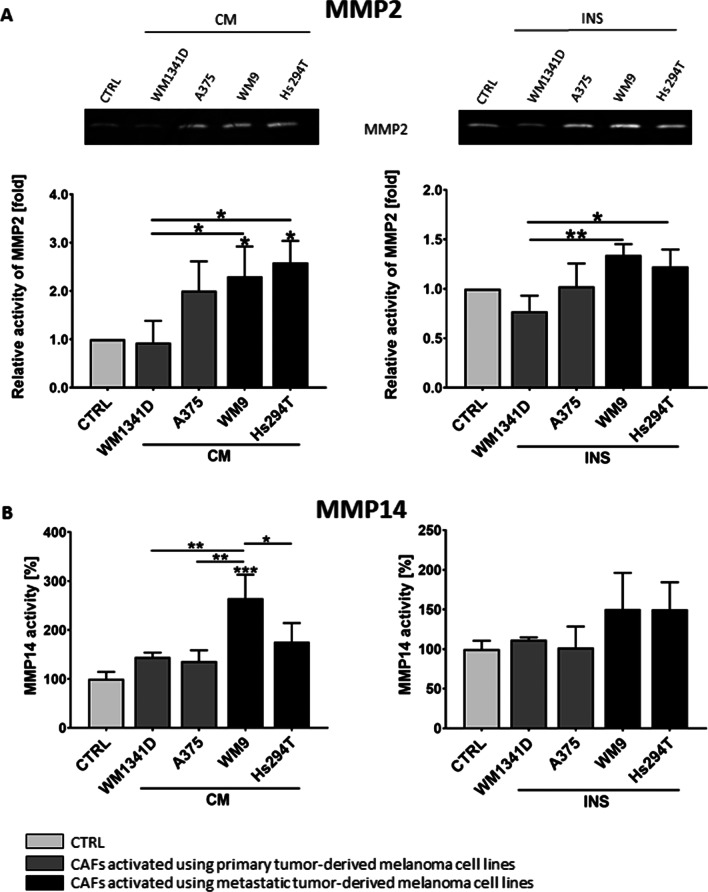
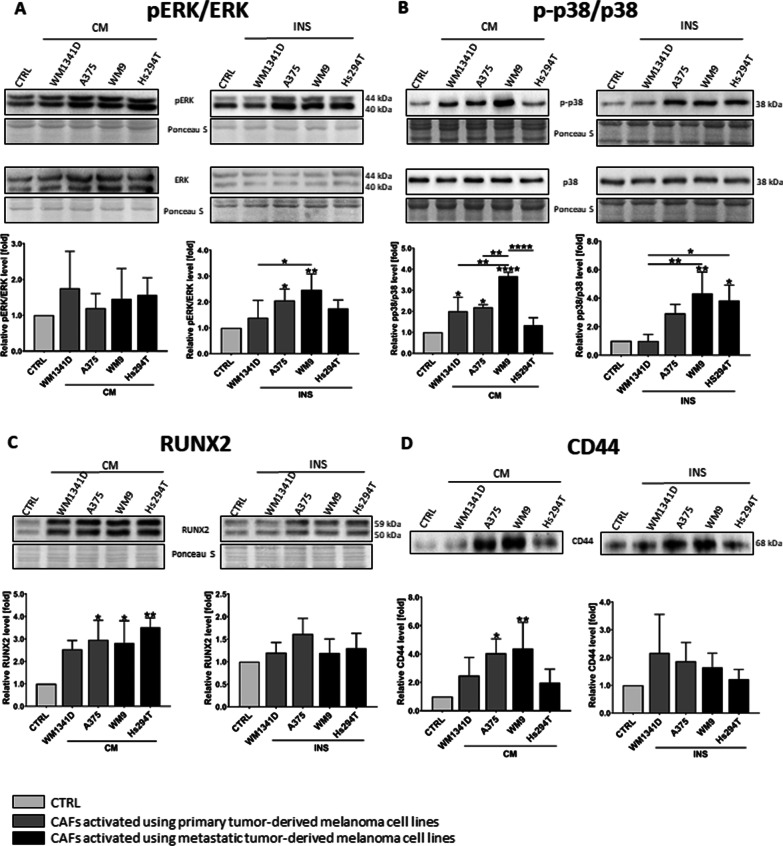
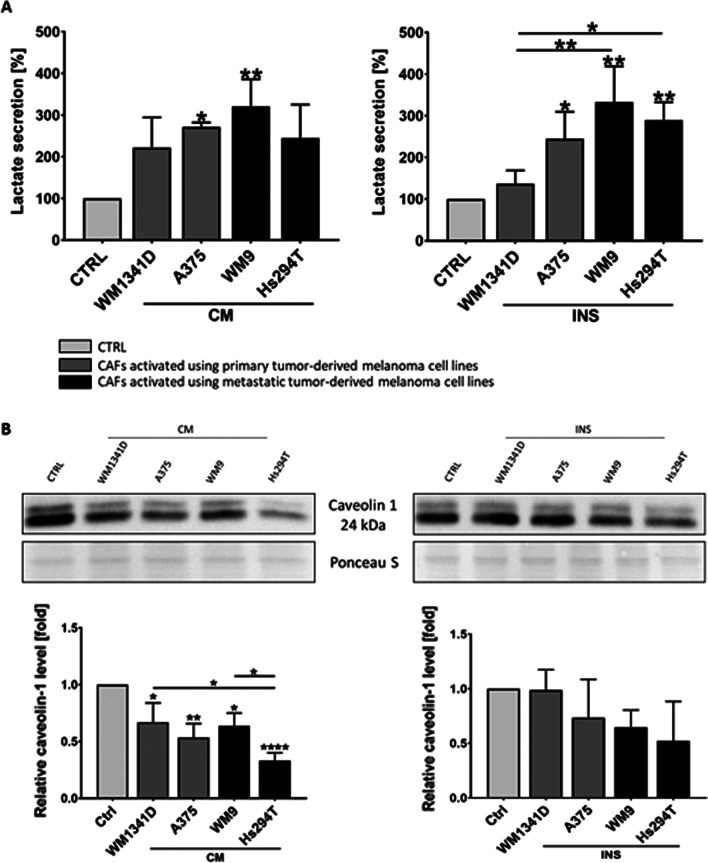
Results: Acquired FAP-α/IL6-positive CAFs exhibited elevated motility expressed as increased migration and invasion ratio, as well as higher proteolytic activity (area of digestion, MMP2, MMP14). Furthermore, fibroblasts activated by melanoma cells showed upregulation of the MMPs' expression mediators' levels (pERK, p-p38, CD44, RUNX), enhanced secretion of lactate, several cytokines (IL8, IL6, CXCL1, CCL2, ICAM1), and proteins related to angiogenesis (GM-CSF, DPPIV, VEGFA, PIGF).
Conclusions: Observed changes in CAFs' biology were mainly driven by highly aggressive melanoma cells (A375, WM9, Hs294T) compared to the less aggressive WM1341D cells and could promote melanoma invasion, as well as impact inflammation, angiogenesis, and acidification of the tumor niche. Interestingly, different approaches to CAFs acquisition seem to complement each other showing interactions between studied cells. Video Abstract.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
