

Mitochondrial antiviral-signalling protein is a client of the BAG6 protein quality control complex
- PMID: 35543156
- PMCID: PMC9264363
- DOI: 10.1242/jcs.259596
Mitochondrial antiviral-signalling protein is a client of the BAG6 protein quality control complex
Abstract
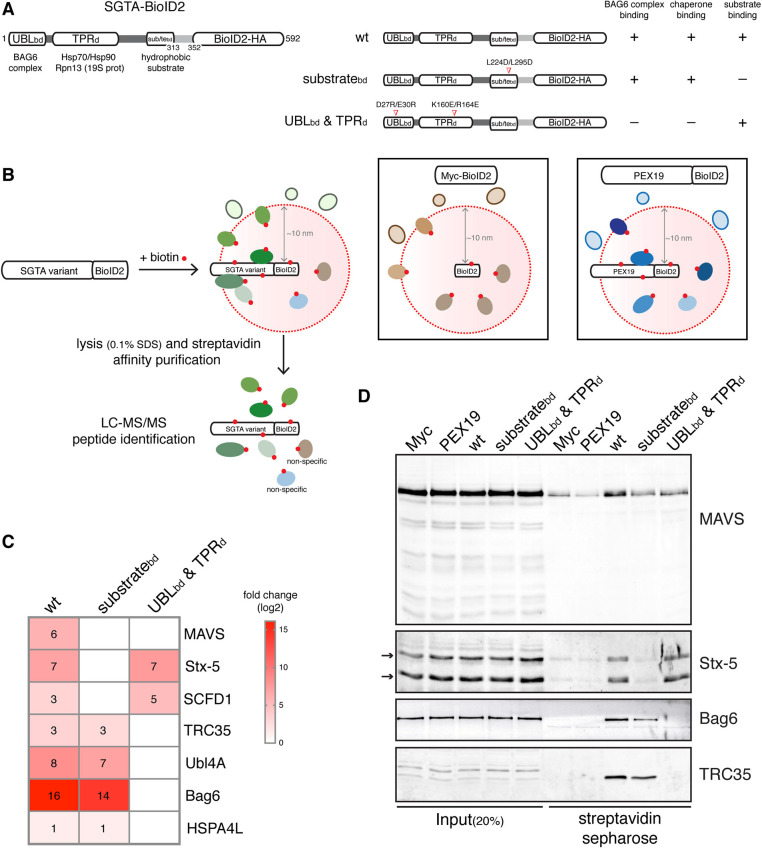
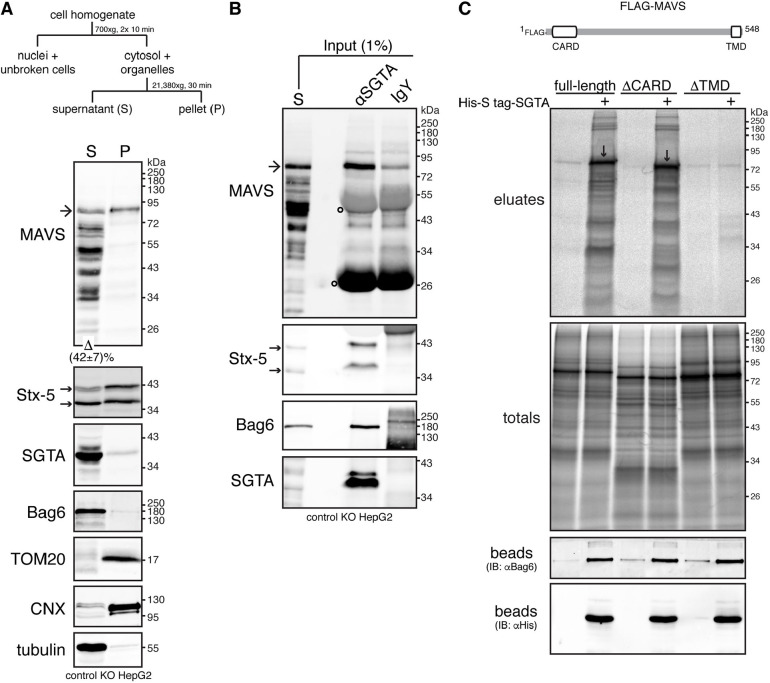
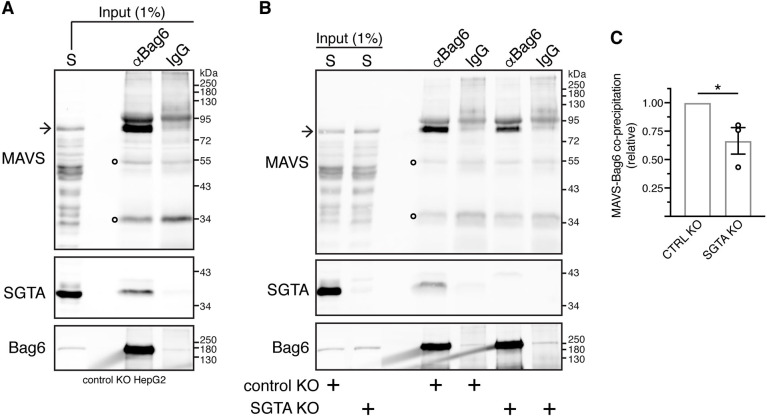
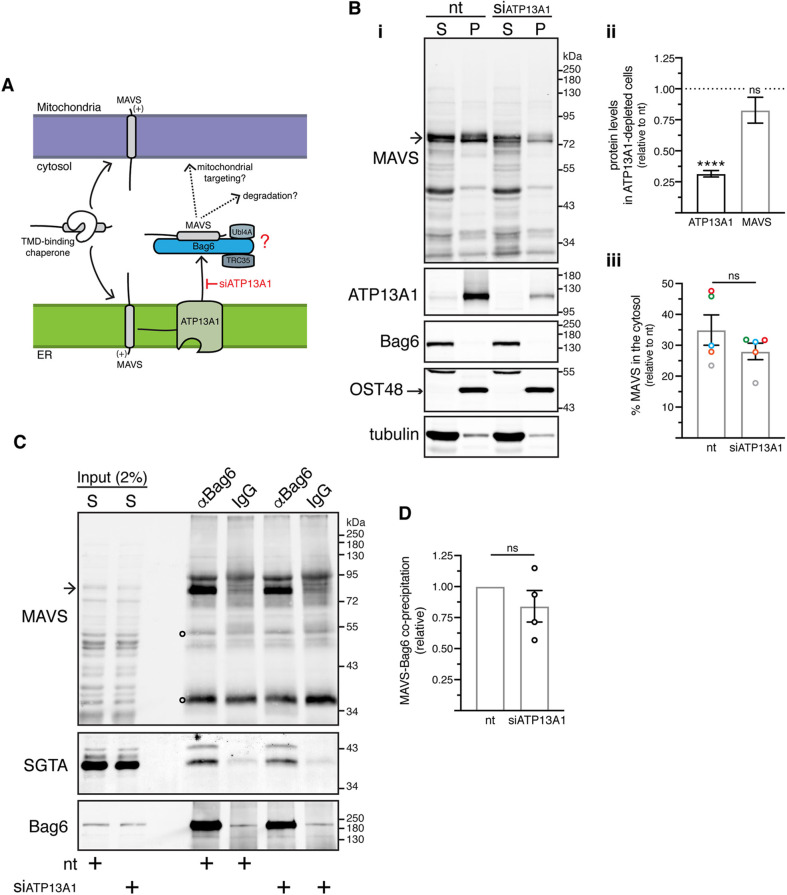
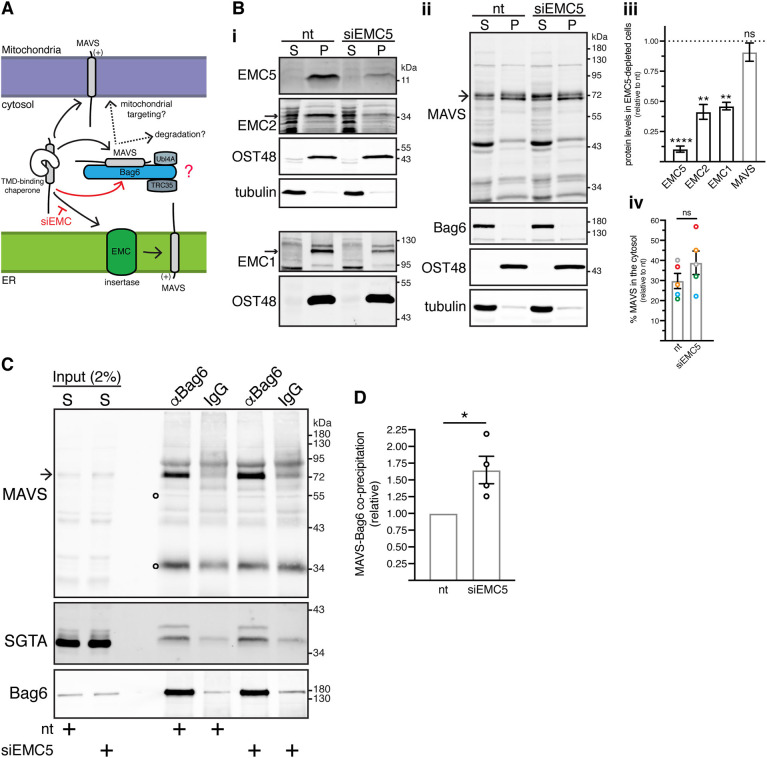
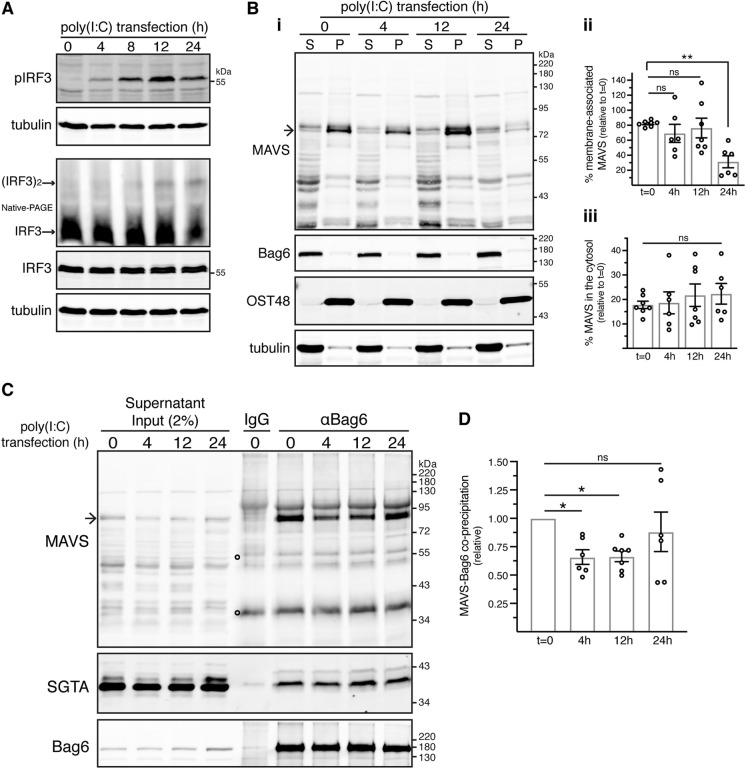
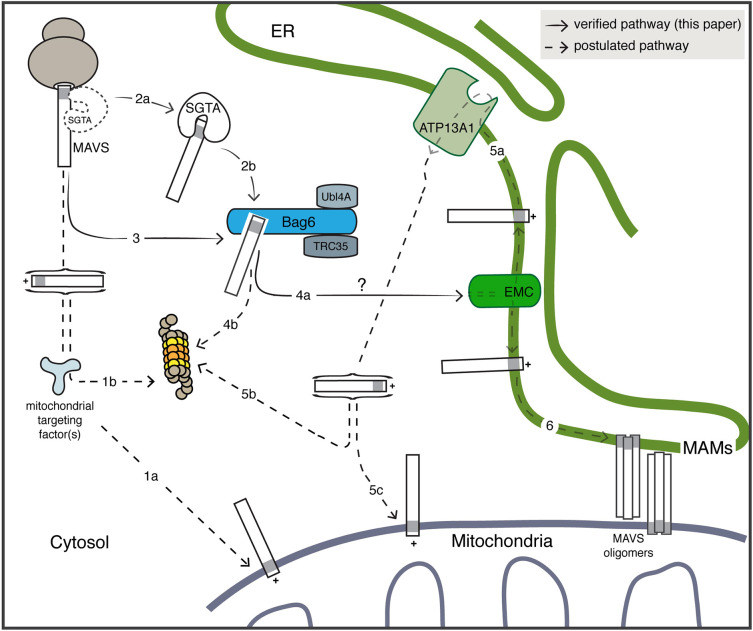
The heterotrimeric BAG6 complex coordinates the direct handover of newly synthesised tail-anchored (TA) membrane proteins from an SGTA-bound preloading complex to the endoplasmic reticulum (ER) delivery component TRC40. In contrast, defective precursors, including aberrant TA proteins, form a stable complex with this cytosolic protein quality control factor, enabling such clients to be either productively re-routed or selectively degraded. We identify the mitochondrial antiviral-signalling protein (MAVS) as an endogenous TA client of both SGTA and the BAG6 complex. Our data suggest that the BAG6 complex binds to a cytosolic pool of MAVS before its misinsertion into the ER membrane, from where it can subsequently be removed via ATP13A1-mediated dislocation. This BAG6-associated fraction of MAVS is dynamic and responds to the activation of an innate immune response, suggesting that BAG6 may modulate the pool of MAVS that is available for coordinating the cellular response to viral infection.
Keywords: BioID2; ER membrane complex; Protein targeting; SGTA; Tail-anchored proteins.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests The authors declare no competing or financial interests.
Figures
Similar articles
-
SGTA antagonizes BAG6-mediated protein triage.Proc Natl Acad Sci U S A. 2012 Nov 20;109(47):19214-9. doi: 10.1073/pnas.1209997109. Epub 2012 Nov 5. Proc Natl Acad Sci U S A. 2012. PMID: 23129660 Free PMC article.
-
SGTA recognizes a noncanonical ubiquitin-like domain in the Bag6-Ubl4A-Trc35 complex to promote endoplasmic reticulum-associated degradation.Cell Rep. 2012 Dec 27;2(6):1633-44. doi: 10.1016/j.celrep.2012.11.010. Epub 2012 Dec 13. Cell Rep. 2012. PMID: 23246001 Free PMC article.
-
The association of BAG6 with SGTA and tail-anchored proteins.PLoS One. 2013;8(3):e59590. doi: 10.1371/journal.pone.0059590. Epub 2013 Mar 22. PLoS One. 2013. PMID: 23533635 Free PMC article.
-
The roles of cytosolic quality control proteins, SGTA and the BAG6 complex, in disease.Adv Protein Chem Struct Biol. 2019;114:265-313. doi: 10.1016/bs.apcsb.2018.11.002. Epub 2018 Dec 18. Adv Protein Chem Struct Biol. 2019. PMID: 30635083 Free PMC article. Review.
-
On the road to nowhere: cross-talk between post-translational protein targeting and cytosolic quality control.Biochem Soc Trans. 2016 Jun 15;44(3):796-801. doi: 10.1042/BST20160045. Biochem Soc Trans. 2016. PMID: 27284044 Review.
Cited by
-
ATP13A1 prevents ERAD of folding-competent mislocalized and misoriented proteins.Mol Cell. 2022 Nov 17;82(22):4277-4289.e10. doi: 10.1016/j.molcel.2022.09.035. Epub 2022 Oct 24. Mol Cell. 2022. PMID: 36283413 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
