

Multiple roles of endocytosis and autophagy in intracellular remodeling during oocyte-to-embryo transition
- PMID: 35545527
- PMCID: PMC9130481
- DOI: 10.2183/pjab.98.013
Multiple roles of endocytosis and autophagy in intracellular remodeling during oocyte-to-embryo transition
Abstract
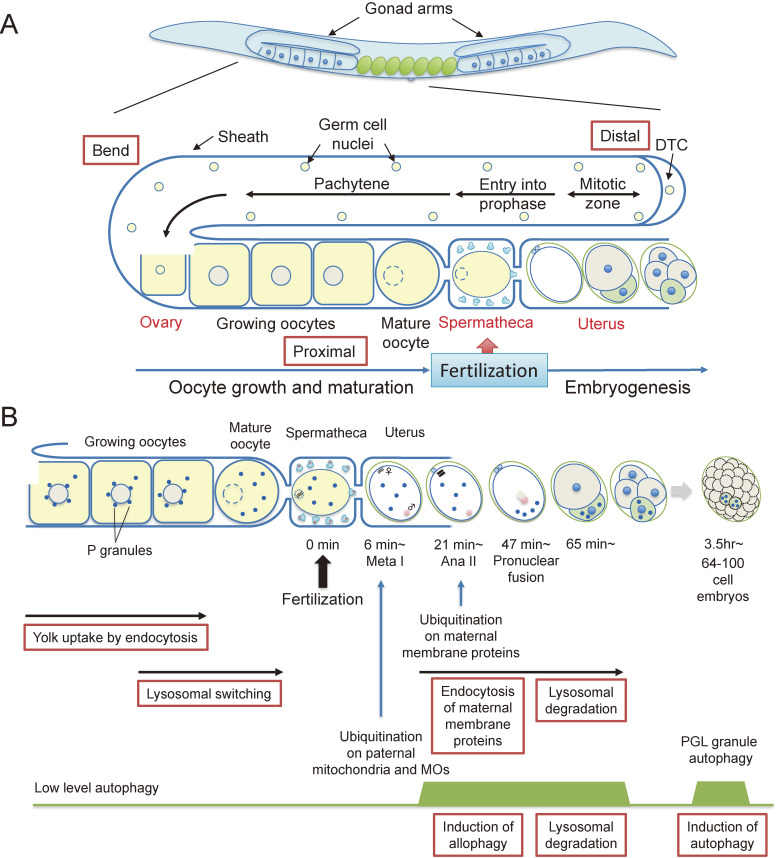
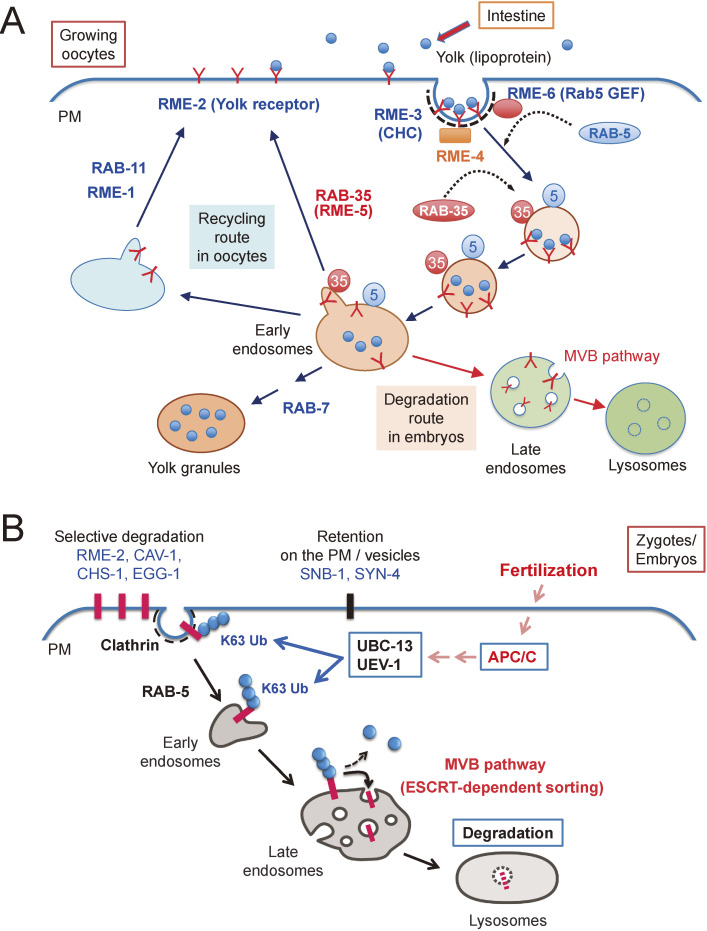
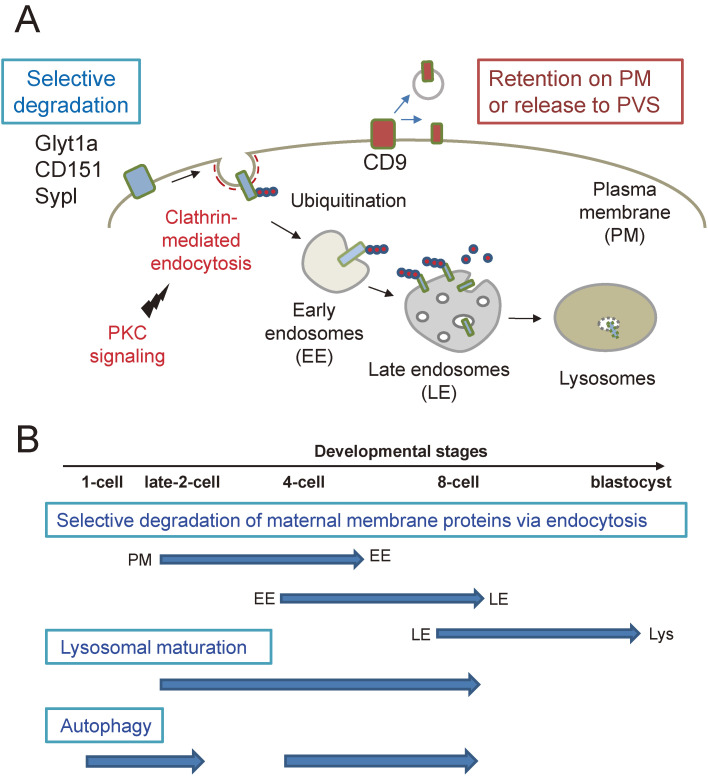
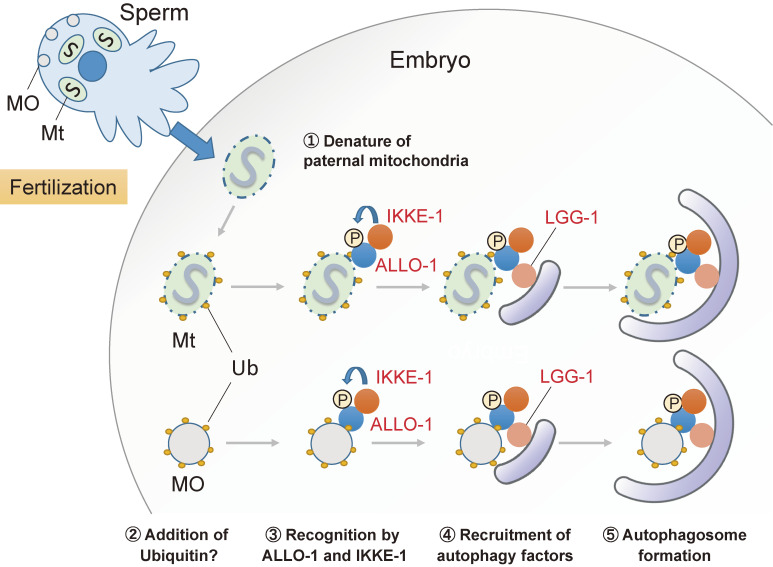
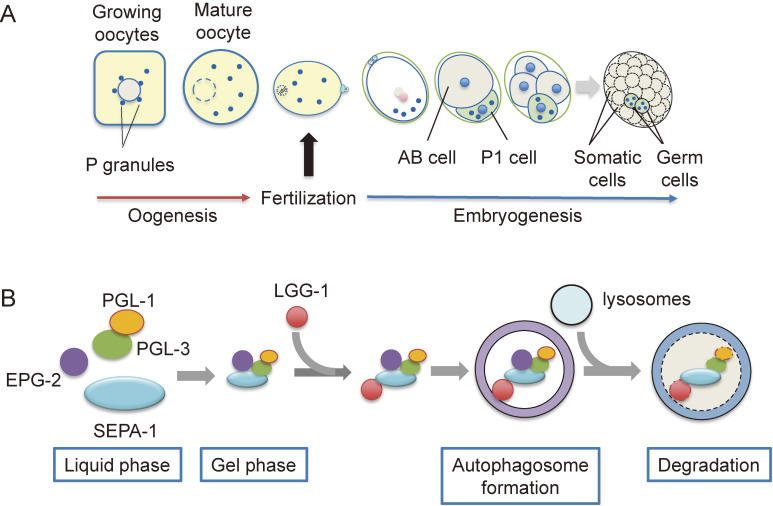
Fertilization is the starting point for creating new progeny. At this time, the highly differentiated oocyte and sperm fuse to form one zygote, which is then converted into a pluripotent early embryo. Recent studies have shown that the lysosomal degradation system via autophagy and endocytosis plays important roles in the remodeling of intracellular components during oocyte-to-embryo transition. For example, in Caenorhabditis elegans, zygotes show high endocytic activity, and some populations of maternal membrane proteins are selectively internalized and delivered to lysosomes for degradation. Furthermore, fertilization triggers selective autophagy of sperm-derived paternal mitochondria, which establishes maternal inheritance of mitochondrial DNA. In addition, it has been shown that autophagy via liquid-liquid phase separation results in the selective degradation of some germ granule components, which are distributed to somatic cells of early embryos. This review outlines the physiological functions of the lysosomal degradation system and its molecular mechanisms in C. elegans and mouse embryos.
Keywords: autophagy; endocytosis; fertilization; oocyte-to-embryo transition.
Figures
Similar articles
-
Degradation of paternal mitochondria by fertilization-triggered autophagy in C. elegans embryos.Science. 2011 Nov 25;334(6059):1141-4. doi: 10.1126/science.1210333. Epub 2011 Oct 13. Science. 2011. PMID: 21998252
-
Maternal inheritance of mitochondrial DNA: degradation of paternal mitochondria by allogeneic organelle autophagy, allophagy.Autophagy. 2012 Mar;8(3):424-5. doi: 10.4161/auto.19243. Epub 2012 Feb 3. Autophagy. 2012. PMID: 22302002
-
Dynamic regulation of autophagy and endocytosis for cell remodeling during early development.Traffic. 2013 May;14(5):479-86. doi: 10.1111/tra.12050. Epub 2013 Feb 19. Traffic. 2013. PMID: 23356349 Review.
-
Dual roles for ubiquitination in the processing of sperm organelles after fertilization.BMC Dev Biol. 2014 Feb 15;14:6. doi: 10.1186/1471-213X-14-6. BMC Dev Biol. 2014. PMID: 24528894 Free PMC article.
-
Autophagosomal Sperm Organelle Clearance and mtDNA Inheritance in C. elegans.Adv Anat Embryol Cell Biol. 2019;231:1-23. doi: 10.1007/102_2018_1. Adv Anat Embryol Cell Biol. 2019. PMID: 30467692 Review.
Cited by
-
MARC-3, a membrane-associated ubiquitin ligase, is required for fast polyspermy block in Caenorhabditis elegans.Nat Commun. 2024 Jan 26;15(1):792. doi: 10.1038/s41467-024-44928-6. Nat Commun. 2024. PMID: 38278786 Free PMC article.
-
Reorganization, specialization, and degradation of oocyte maternal components for early development.Reprod Med Biol. 2023 Jan 28;22(1):e12505. doi: 10.1002/rmb2.12505. eCollection 2023 Jan-Dec. Reprod Med Biol. 2023. PMID: 36726596 Free PMC article. Review.
-
Mitochondrial Deoxyribonucleic Acid (mtDNA), Maternal Inheritance, and Their Role in the Development of Cancers: A Scoping Review.Cureus. 2023 Jun 1;15(6):e39812. doi: 10.7759/cureus.39812. eCollection 2023 Jun. Cureus. 2023. PMID: 37397663 Free PMC article.
-
VPS34 Governs Oocyte Developmental Competence by Regulating Mito/Autophagy: A Novel Insight into the Significance of RAB7 Activity and Its Subcellular Location.Adv Sci (Weinh). 2024 Nov;11(41):e2308823. doi: 10.1002/advs.202308823. Epub 2024 Sep 17. Adv Sci (Weinh). 2024. PMID: 39287146 Free PMC article.
-
Endosomal-lysosomal organellar assembly (ELYSA) structures coordinate lysosomal degradation systems through mammalian oocyte-to-embryo transition.Elife. 2025 Mar 17;13:RP99358. doi: 10.7554/eLife.99358. Elife. 2025. PMID: 40094202 Free PMC article.
References
-
- Tadros W., Lipshitz H.D. (2005) Setting the stage for development: mRNA translation and stability during oocyte maturation and egg activation in Drosophila. Dev. Dyn. 232, 593–608. - PubMed
-
- Schier A.F. (2007) The maternal-zygotic transition: death and birth of RNAs. Science 316, 406–407. - PubMed
-
- Merz E.A., Brinster R.L., Brunner S., Chen H.Y. (1981) Protein degradation during preimplantation development of the mouse. J. Reprod. Fertil. 61, 415–418. - PubMed
-
- Marcello M.R., Singson A. (2010) Fertilization and the oocyte-to-embryo transition in C. elegans. BMB Rep. 43, 389–399. - PubMed
-
- Miller M.A., Nguyen V.Q., Lee M.H., Kosinski M., Schedl T., Caprioli R.M., et al. (2001) A sperm cytoskeletal protein that signals oocyte meiotic maturation and ovulation. Science 291, 2144–2147. - PubMed