

High-fat diet-induced diabetes couples to Alzheimer's disease through inflammation-activated C/EBPβ/AEP pathway
- PMID: 35546632
- PMCID: PMC10032575
- DOI: 10.1038/s41380-022-01600-z
High-fat diet-induced diabetes couples to Alzheimer's disease through inflammation-activated C/EBPβ/AEP pathway
Abstract
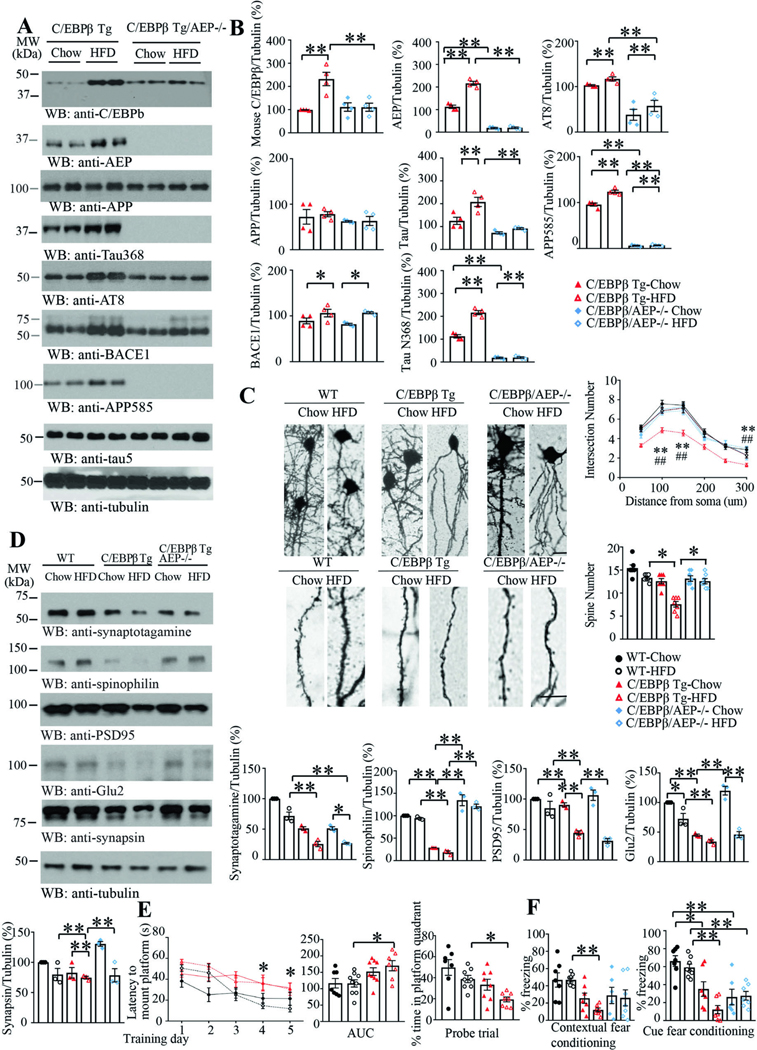
Diabetes is a risk factor for Alzheimer's disease (AD), which is also called type 3 diabetes with insulin reduction and insulin resistance in AD patient brains. However, the molecular mechanism coupling diabetes to AD onset remains incompletely understood. Here we show that inflammation, associated with obesity and diabetes elicited by high-fat diet (HFD), activates neuronal C/EBPβ/AEP signaling that drives AD pathologies and cognitive disorders. HFD stimulates diabetes and insulin resistance in neuronal Thy1-C/EBPβ transgenic (Tg) mice, accompanied with prominent mouse Aβ accumulation and hyperphosphorylated Tau aggregation in the brain, triggering cognitive deficits. These effects are profoundly diminished when AEP is deleted from C/EBPβ Tg mice. Chronic treatment with inflammatory lipopolysaccharide (LPS) facilitates AD pathologies and cognitive disorders in C/EBPβ Tg but not in wild-type mice, and these deleterious effects were substantially alleviated in C/EBPβ Tg/AEP -/- mice. Remarkably, the anti-inflammatory drug aspirin strongly attenuates HFD-induced diabetes and AD pathologies in neuronal C/EBPβ Tg mice. Therefore, our findings demonstrate that inflammation-activated neuronal C/EBPβ/AEP signaling couples diabetes to AD.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
COMPETING INTERESTS
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
