

The Hidden Side of Complement Regulator C4BP: Dissection and Evaluation of Its Immunomodulatory Activity
- PMID: 35547734
- PMCID: PMC9084231
- DOI: 10.3389/fimmu.2022.883743
The Hidden Side of Complement Regulator C4BP: Dissection and Evaluation of Its Immunomodulatory Activity
Abstract
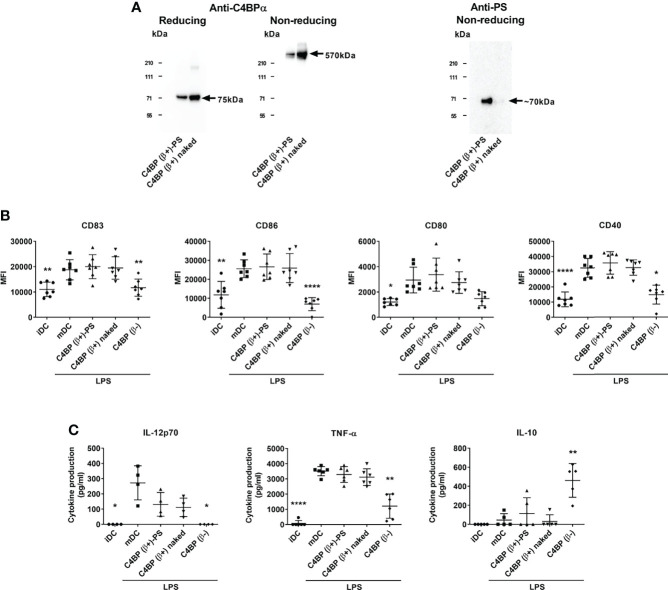
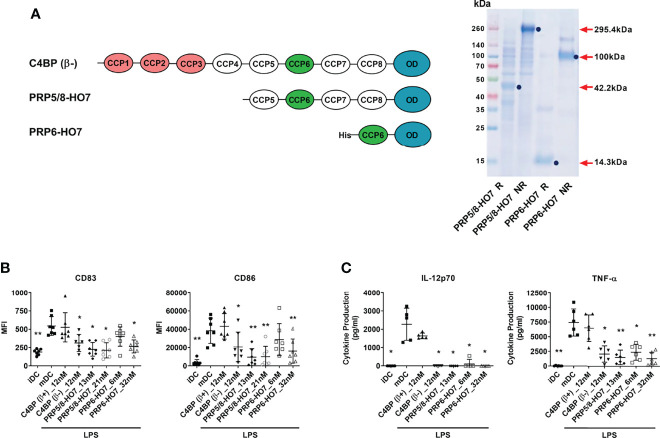
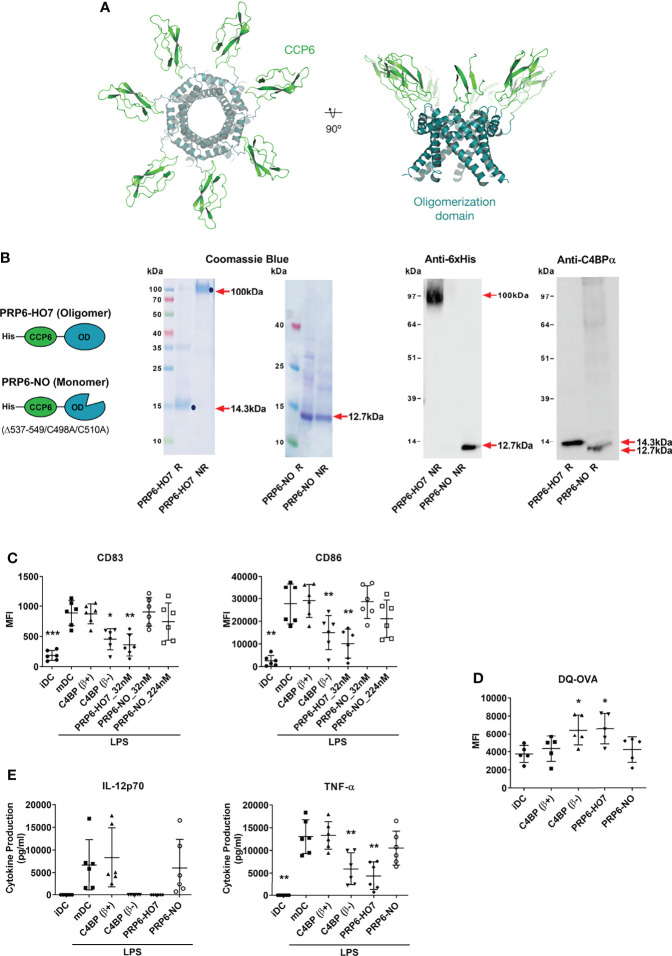
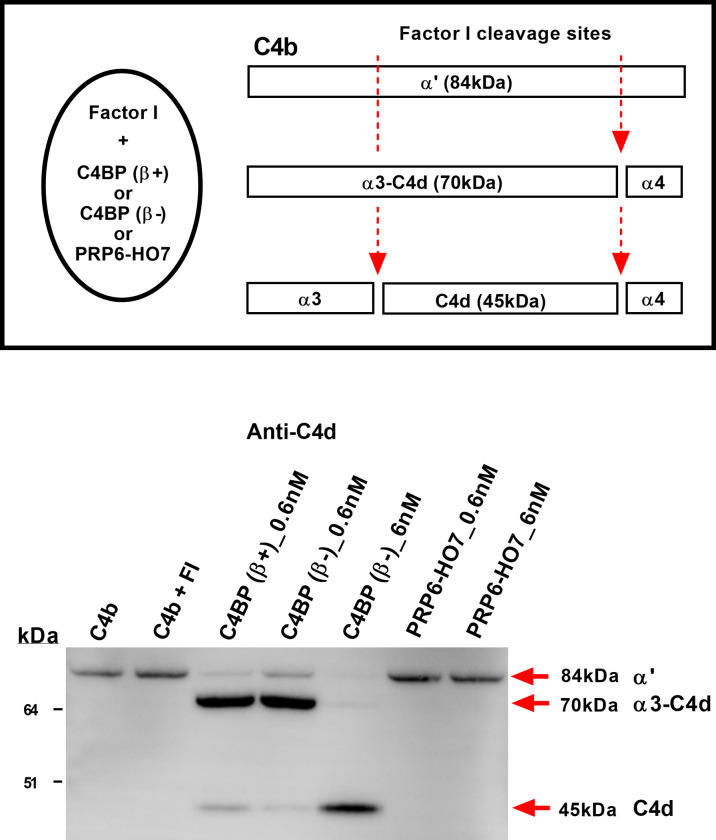
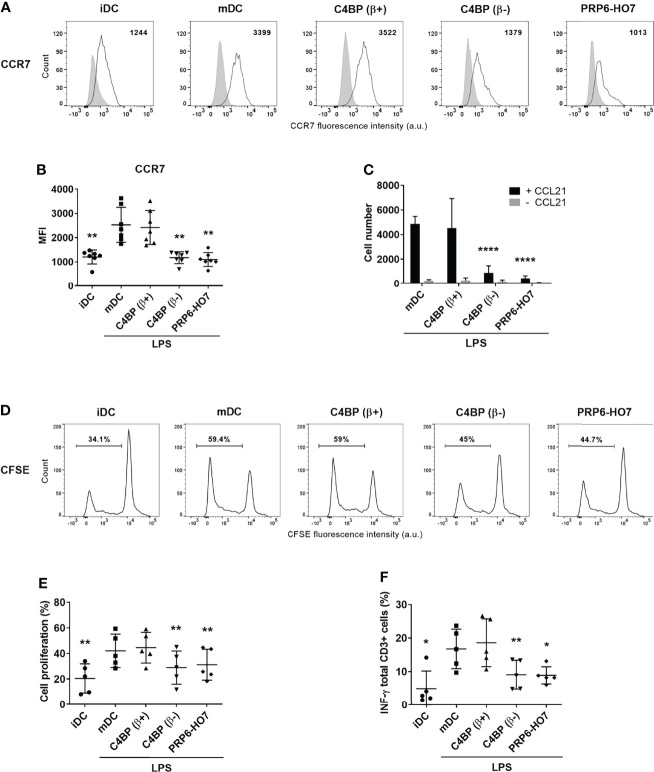
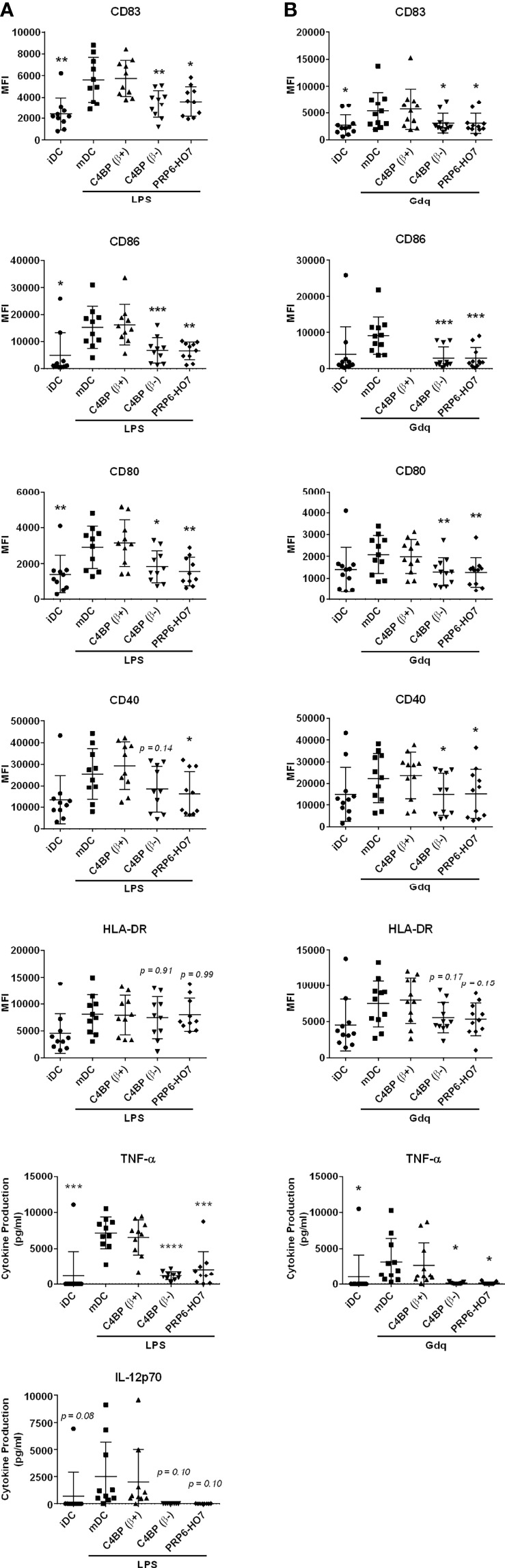
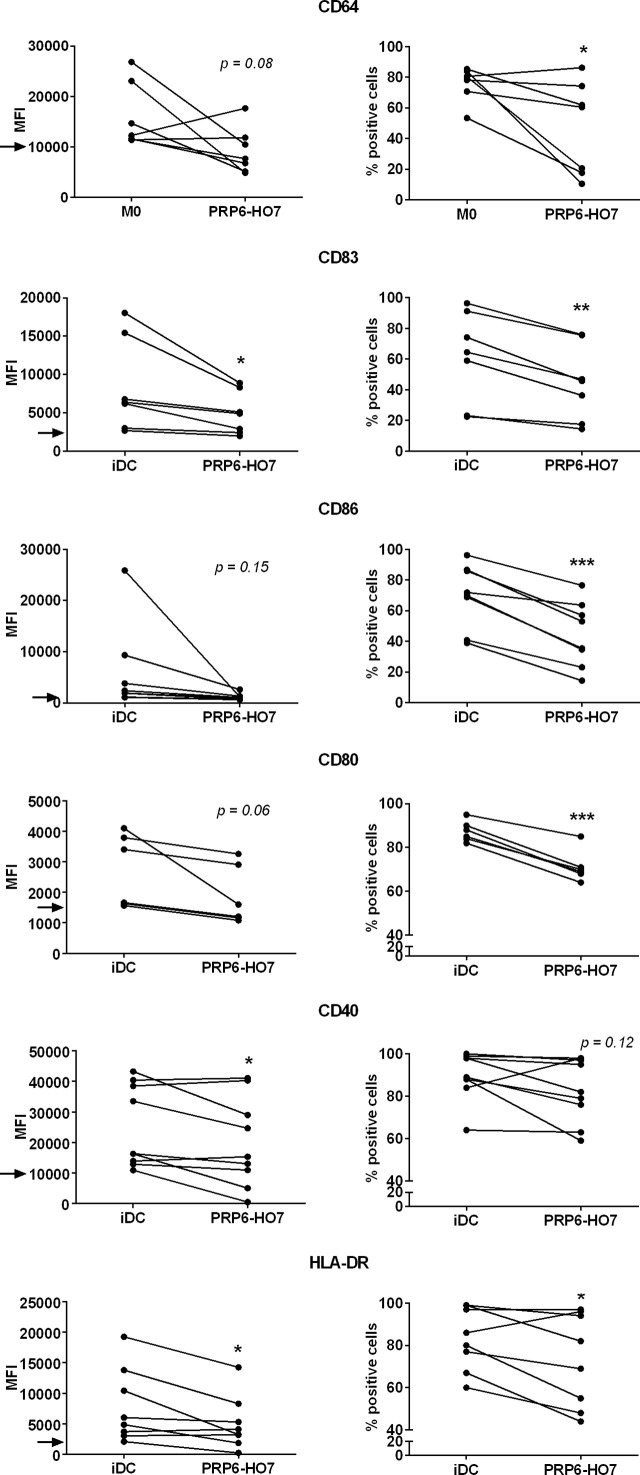
C4b-binding protein (C4BP) is a well-known regulator of the complement system that holds additional and important activities unrelated to complement inhibition. Recently, we have described a novel immunomodulatory activity in the minor C4BP(β-) isoform directly acting over inflammatory phagocytes. Here we show that incorporation of the β-chain to the C4BP α-chain oligomer interferes with this immunomodulatory activity of C4BP. Moreover, an oligomeric form including only the complement control protein 6 (CCP6) domain of the C4BP α-chain (PRP6-HO7) is sufficient to "reprogram" monocyte-derived DCs (Mo-DCs) from a pro-inflammatory and immunogenic phenotype to an anti-inflammatory and tolerogenic state. PRP6-HO7 lacks complement regulatory activity but retains full immunomodulatory activity over inflammatory Mo-DCs induced by TLRs, characterized by downregulation of relevant surface markers such as CD83, HLA-DR, co-stimulatory molecules such as CD86, CD80 and CD40, and pro-inflammatory cytokines such as IL-12 and TNF-α. Furthermore, PRP6-HO7-treated Mo-DCs shows increased endocytosis, significantly reduced CCR7 expression and CCL21-mediated chemotaxis, and prevents T cell alloproliferation. Finally, PRP6-HO7 shows also full immunomodulatory activity over Mo-DCs isolated from lupus nephritis patients with active disease, even without further pro-inflammatory stimulation. Therefore PRP6-HO7, retaining the immunomodulatory activity of C4BP(β-) and lacking its complement regulatory activity, might represent a promising and novel alternative to treat autoimmune diseases.
Keywords: C4BP(β-); PRP6-HO7; dendritic cells; immunomodulation; inflammation; lupus nephritis.
Copyright © 2022 Serrano, Luque, Mitjavila, Blom, Rodríguez de Córdoba, Vega, Torras and Aran.
Conflict of interest statement
IS, AL and JA are co-inventors on pending or issued patents involving compounds and methods for immunomodulation. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


Similar articles
-
C4BP(β-)-mediated immunomodulation attenuates inflammation in DSS-induced murine colitis and in myeloid cells from IBD patients.Pharmacol Res. 2023 Nov;197:106948. doi: 10.1016/j.phrs.2023.106948. Epub 2023 Oct 6. Pharmacol Res. 2023. PMID: 37806602
-
The α7β0 isoform of the complement regulator C4b-binding protein induces a semimature, anti-inflammatory state in dendritic cells.J Immunol. 2013 Mar 15;190(6):2857-72. doi: 10.4049/jimmunol.1200503. Epub 2013 Feb 6. J Immunol. 2013. PMID: 23390292
-
The Complement Inhibitor Factor H Generates an Anti-Inflammatory and Tolerogenic State in Monocyte-Derived Dendritic Cells.J Immunol. 2016 May 15;196(10):4274-90. doi: 10.4049/jimmunol.1500455. Epub 2016 Apr 13. J Immunol. 2016. PMID: 27076676
-
C4b-binding protein: The good, the bad and the deadly. Novel functions of an old friend.Immunol Lett. 2016 Jan;169:82-92. doi: 10.1016/j.imlet.2015.11.014. Epub 2015 Dec 2. Immunol Lett. 2016. PMID: 26658464 Review.
-
C4b-binding protein: a forgotten factor in thrombosis and hemostasis.Semin Thromb Hemost. 2011 Jun;37(4):355-61. doi: 10.1055/s-0031-1276584. Epub 2011 Jul 30. Semin Thromb Hemost. 2011. PMID: 21805441 Review.
Cited by
-
Smoking Impacts Alzheimer's Disease Progression Through Oral Microbiota Modulation.Mol Neurobiol. 2025 Jan;62(1):19-44. doi: 10.1007/s12035-024-04241-1. Epub 2024 May 25. Mol Neurobiol. 2025. PMID: 38795302 Review.
-
Neisseria gonorrhoeae co-opts C4b-binding protein to enhance complement-independent survival from neutrophils.PLoS Pathog. 2023 Mar 2;19(3):e1011055. doi: 10.1371/journal.ppat.1011055. eCollection 2023 Mar. PLoS Pathog. 2023. PMID: 36862761 Free PMC article.
-
Diverse Functions of C4b-Binding Protein in Health and Disease.J Immunol. 2023 Nov 15;211(10):1443-1449. doi: 10.4049/jimmunol.2300333. J Immunol. 2023. PMID: 37931209 Free PMC article. Review.
-
C4b-binding protein inhibits particulate- and crystalline-induced NLRP3 inflammasome activation.Front Immunol. 2023 May 22;14:1149822. doi: 10.3389/fimmu.2023.1149822. eCollection 2023. Front Immunol. 2023. PMID: 37283747 Free PMC article.
-
Screening of Exosome-Derived Proteins and Their Potential as Biomarkers in Diagnostic and Prognostic for Pancreatic Cancer.Int J Mol Sci. 2023 Aug 9;24(16):12604. doi: 10.3390/ijms241612604. Int J Mol Sci. 2023. PMID: 37628784 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous