

Monitoring Autophagy in Rice With GFP-ATG8 Marker Lines
- PMID: 35548298
- PMCID: PMC9083259
- DOI: 10.3389/fpls.2022.866367
Monitoring Autophagy in Rice With GFP-ATG8 Marker Lines
Abstract
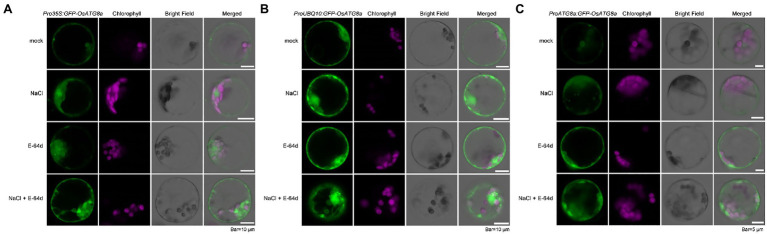
Autophagy is a conserved intracellular trafficking pathway for bulk degradation and recycling of cellular components in eukaryotes. The hallmark of autophagy is the formation of double-membraned vesicles termed autophagosomes, which selectively or non-selectively pack up various macromolecules and organelles and deliver these cargoes into the vacuole/lysosome. Like all other membrane trafficking pathways, the observation of autophagy is largely dependent on marker lines. ATG8/LC3 is the only autophagy-related (ATG) protein that, through a covalent bond to phosphatidylethanolamine (PE), associates tightly with the isolation membrane/pre-autophagosomal structure (PAS), the growing phagophore, the mature autophagosome, and the autophagic bodies. Therefore, fluorescent protein (FP)-tagged ATG8 had been widely used for monitoring autophagosome formation and autophagic flux. In rice (Oryza sativa), FP-OsATG8 driven by Cauliflower mosaic virus (CaMV) 35S promoter had been used for imaging autophagosome and autophagic bodies. Here, we constructed three vectors carrying GFP-OsATG8a, driven by 35S, ubiquitin, and the endogenous ATG8a promoter, individually. Then, we compared them for their suitability in monitoring autophagy, by observing GFP-ATG8a puncta formation in transiently transformed rice protoplasts, and by tracking the autophagic flux with GFP-ATG8 cleavage assay in rice stable transgenic lines. GFP-Trap immunoprecipitation and mass spectrometry were also performed with the three marker lines to show that they can be used reliably for proteomic studies. We found out that the ubiquitin promoter is the best for protoplast imaging. Transgenic rice seedlings of the three marker lines showed comparable performance in autophagic flux measurement using the GFP-ATG8 cleavage assay. Surprisingly, the levels of GFP-ATG8a transcripts and protein contents were similar in all marker lines, indicating post-transcriptional regulation of the transgene expression by a yet unknown mechanism. These marker lines can serve as useful tools for autophagy studies in rice.
Keywords: ATG8; autophagic flux; autophagy; post-transcriptional regulation; rice.
Copyright © 2022 Liu, Zhang, Yang, Liu and Gong.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




Similar articles
-
Visualization of Atg3 during autophagosome formation in Saccharomyces cerevisiae.J Biol Chem. 2015 Mar 27;290(13):8146-53. doi: 10.1074/jbc.M114.626952. Epub 2015 Feb 2. J Biol Chem. 2015. PMID: 25645919 Free PMC article.
-
The two Dictyostelium discoideum autophagy 8 proteins have distinct autophagic functions.Eur J Cell Biol. 2017 Jun;96(4):312-324. doi: 10.1016/j.ejcb.2017.03.014. Epub 2017 Apr 4. Eur J Cell Biol. 2017. PMID: 28413119
-
Analysis of Autophagy Under Abiotic Stress in Arabidopsis Seedlings Expressing the GFP-ATG8 Autophagosome Marker.Curr Protoc. 2025 Jun;5(6):e70166. doi: 10.1002/cpz1.70166. Curr Protoc. 2025. PMID: 40557747 Free PMC article.
-
Molecular Mechanisms of Autophagy in Plants: Role of ATG8 Proteins in Formation and Functioning of Autophagosomes.Biochemistry (Mosc). 2016 Apr;81(4):348-63. doi: 10.1134/S0006297916040052. Biochemistry (Mosc). 2016. PMID: 27293092 Review.
-
Non-canonical roles of ATG8 for TFEB activation.Biochem Soc Trans. 2022 Feb 28;50(1):47-54. doi: 10.1042/BST20210813. Biochem Soc Trans. 2022. PMID: 35166325 Review.
Cited by
-
Recruitment of Atg1 to the phagophore by Atg8 orchestrates autophagy machineries.Nat Struct Mol Biol. 2025 Apr 28. doi: 10.1038/s41594-025-01546-0. Online ahead of print. Nat Struct Mol Biol. 2025. PMID: 40295771
-
Myotubularin 2 interacts with SEC23A and negatively regulates autophagy at ER exit sites in Arabidopsis.Autophagy. 2025 Jan;21(1):141-159. doi: 10.1080/15548627.2024.2394302. Epub 2024 Sep 8. Autophagy. 2025. PMID: 39177202 Free PMC article.
-
Activation of autophagy by Citri Reticulatae Semen extract ameliorates amyloid-beta-induced cell death and cognition deficits in Alzheimer's disease.Neural Regen Res. 2024 Nov 1;19(11):2467-2479. doi: 10.4103/NRR.NRR-D-23-00954. Epub 2024 Jan 31. Neural Regen Res. 2024. PMID: 38526283 Free PMC article.
-
Interaction of Potato Autophagy-Related StATG8 Family Proteins with Pathogen Effector and WRKY Transcription Factor in the Nucleus.Microorganisms. 2025 Jul 5;13(7):1589. doi: 10.3390/microorganisms13071589. Microorganisms. 2025. PMID: 40732098 Free PMC article.
-
Studying plant autophagy: challenges and recommended methodologies.Adv Biotechnol (Singap). 2023 Oct 26;1(4):2. doi: 10.1007/s44307-023-00002-8. Adv Biotechnol (Singap). 2023. PMID: 39883189 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources