

Functional and Structural Diversity of Bacterial Contact-Dependent Growth Inhibition Effectors
- PMID: 35558562
- PMCID: PMC9086364
- DOI: 10.3389/fmolb.2022.866854
Functional and Structural Diversity of Bacterial Contact-Dependent Growth Inhibition Effectors
Abstract
Bacteria live in complex communities and environments, competing for space and nutrients. Within their niche habitats, bacteria have developed various inter-bacterial mechanisms to compete and communicate. One such mechanism is contact-dependent growth inhibition (CDI). CDI is found in many Gram-negative bacteria, including several pathogens. These CDI+ bacteria encode a CdiB/CdiA two-partner secretion system that delivers inhibitory toxins into neighboring cells upon contact. Toxin translocation results in the growth inhibition of closely related strains and provides a competitive advantage to the CDI+ bacteria. CdiB, an outer-membrane protein, secretes CdiA onto the surface of the CDI+ bacteria. When CdiA interacts with specific target-cell receptors, CdiA delivers its C-terminal toxin region (CdiA-CT) into the target-cell. CdiA-CT toxin proteins display a diverse range of toxic functions, such as DNase, RNase, or pore-forming toxin activity. CDI+ bacteria also encode an immunity protein, CdiI, that specifically binds and neutralizes its cognate CdiA-CT, protecting the CDI+ bacteria from auto-inhibition. In Gram-negative bacteria, toxin/immunity (CdiA-CT/CdiI) pairs have highly variable sequences and functions, with over 130 predicted divergent toxin/immunity complex families. In this review, we will discuss biochemical and structural advances made in the characterization of CDI. This review will focus on the diverse array of CDI toxin/immunity complex structures together with their distinct toxin functions. Additionally, we will discuss the most recent studies on target-cell recognition and toxin entry, along with the discovery of a new member of the CDI loci. Finally, we will offer insights into how these diverse toxin/immunity complexes could be harnessed to fight human diseases.
Keywords: contact-dependent growth inhibition; structure; toxin function; toxin/immunity complex; type V secretion system.
Copyright © 2022 Cuthbert, Hayes and Goulding.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

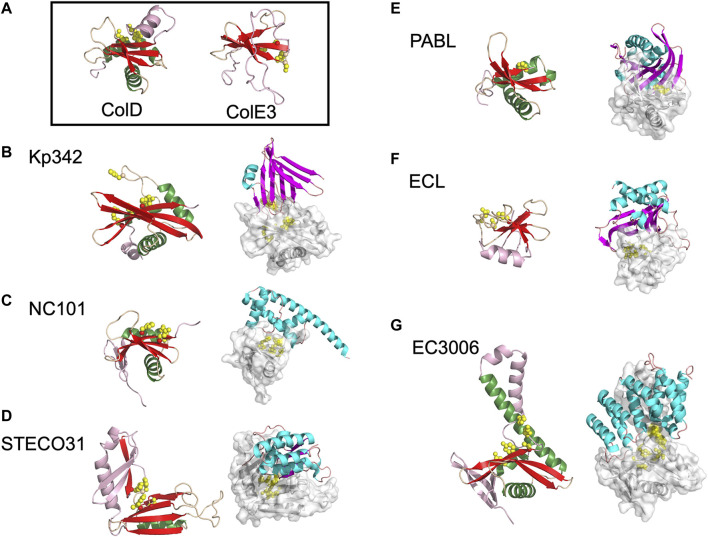
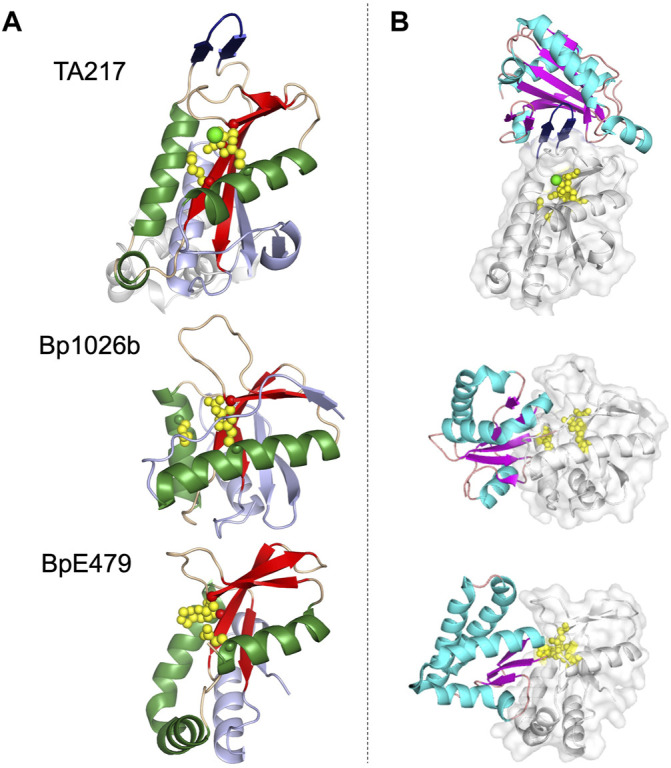
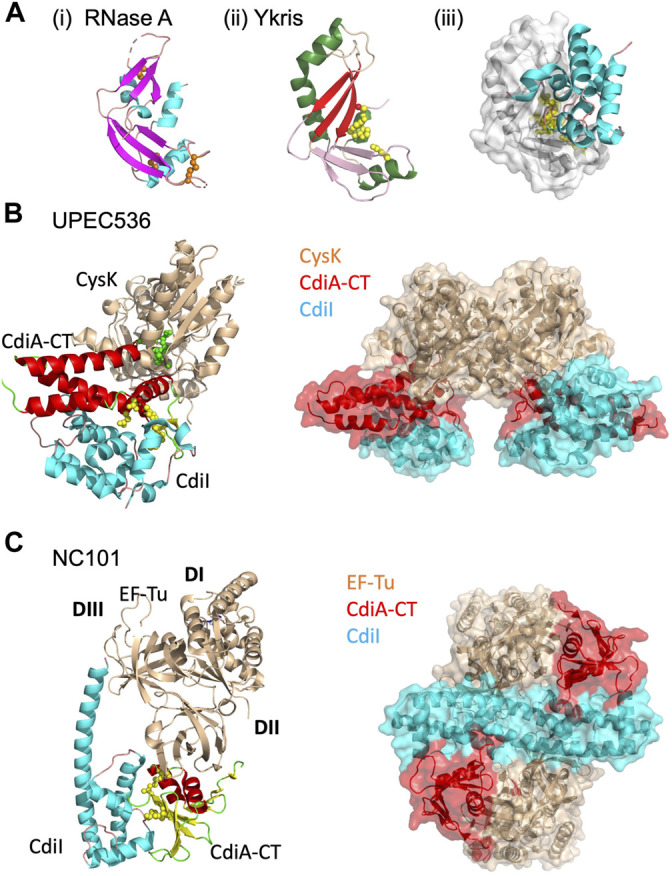
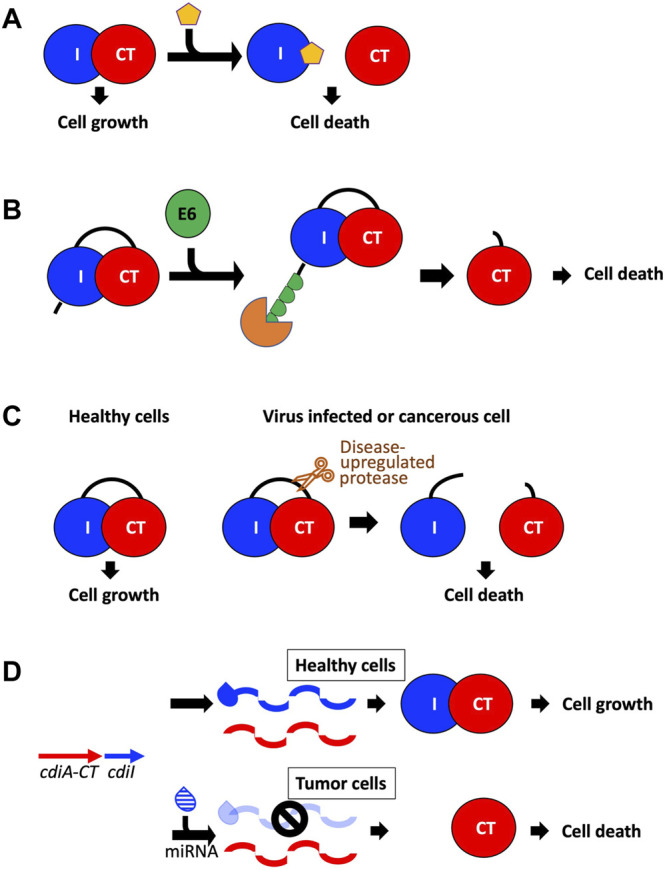
Similar articles
-
Contact-Dependent Growth Inhibition (CDI) and CdiB/CdiA Two-Partner Secretion Proteins.J Mol Biol. 2015 Nov 20;427(23):3754-65. doi: 10.1016/j.jmb.2015.09.010. Epub 2015 Sep 24. J Mol Biol. 2015. PMID: 26388411 Free PMC article. Review.
-
A widespread family of polymorphic contact-dependent toxin delivery systems in bacteria.Nature. 2010 Nov 18;468(7322):439-42. doi: 10.1038/nature09490. Nature. 2010. PMID: 21085179 Free PMC article.
-
Identification of functional toxin/immunity genes linked to contact-dependent growth inhibition (CDI) and rearrangement hotspot (Rhs) systems.PLoS Genet. 2011 Aug;7(8):e1002217. doi: 10.1371/journal.pgen.1002217. Epub 2011 Aug 4. PLoS Genet. 2011. PMID: 21829394 Free PMC article.
-
Diversification of β-Augmentation Interactions between CDI Toxin/Immunity Proteins.J Mol Biol. 2015 Nov 20;427(23):3766-84. doi: 10.1016/j.jmb.2015.09.020. Epub 2015 Oct 9. J Mol Biol. 2015. PMID: 26449640 Free PMC article.
-
Mechanisms and biological roles of contact-dependent growth inhibition systems.Cold Spring Harb Perspect Med. 2014 Feb 1;4(2):a010025. doi: 10.1101/cshperspect.a010025. Cold Spring Harb Perspect Med. 2014. PMID: 24492845 Free PMC article. Review.
Cited by
-
The type II secretion system as an underappreciated and understudied mediator of interbacterial antagonism.Infect Immun. 2024 Aug 13;92(8):e0020724. doi: 10.1128/iai.00207-24. Epub 2024 Jul 9. Infect Immun. 2024. PMID: 38980047 Free PMC article. Review.
-
Mechanism of activation of contact-dependent growth inhibition tRNase toxin by the amino acid biogenesis factor CysK in the bacterial competition system.Nucleic Acids Res. 2025 Jan 7;53(1):gkae735. doi: 10.1093/nar/gkae735. Nucleic Acids Res. 2025. PMID: 39228374 Free PMC article.
-
Effectiveness of Pseudomonas aeruginosa type VI secretion system relies on toxin potency and type IV pili-dependent interaction.PLoS Pathog. 2023 May 30;19(5):e1011428. doi: 10.1371/journal.ppat.1011428. eCollection 2023 May. PLoS Pathog. 2023. PMID: 37253075 Free PMC article.
-
Pangenome Analysis Reveals Novel Contact-Dependent Growth Inhibition System and Phenazine Biosynthesis Operons in Proteus mirabilis BL95 That Are Located in An Integrative and Conjugative Element.Microorganisms. 2024 Jun 28;12(7):1321. doi: 10.3390/microorganisms12071321. Microorganisms. 2024. PMID: 39065090 Free PMC article.
-
Autoproteolytic mechanism of CdiA toxin release reconstituted in vitro.J Bacteriol. 2024 Oct 24;206(10):e0024924. doi: 10.1128/jb.00249-24. Epub 2024 Sep 30. J Bacteriol. 2024. PMID: 39347575 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases