

SNAT7 regulates mTORC1 via macropinocytosis
- PMID: 35561222
- PMCID: PMC9171778
- DOI: 10.1073/pnas.2123261119
SNAT7 regulates mTORC1 via macropinocytosis
Abstract
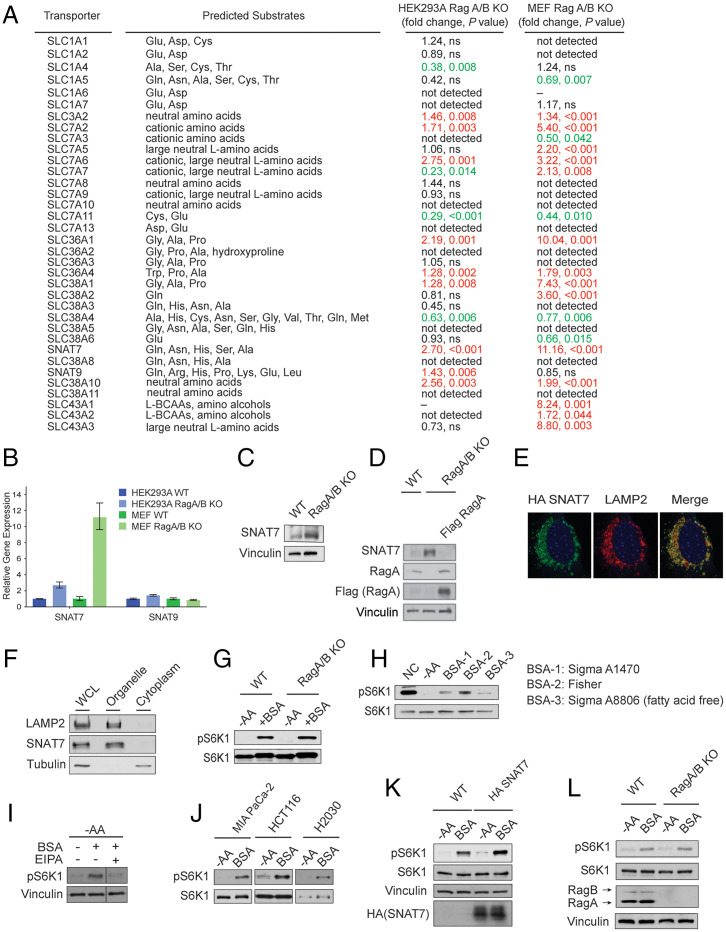
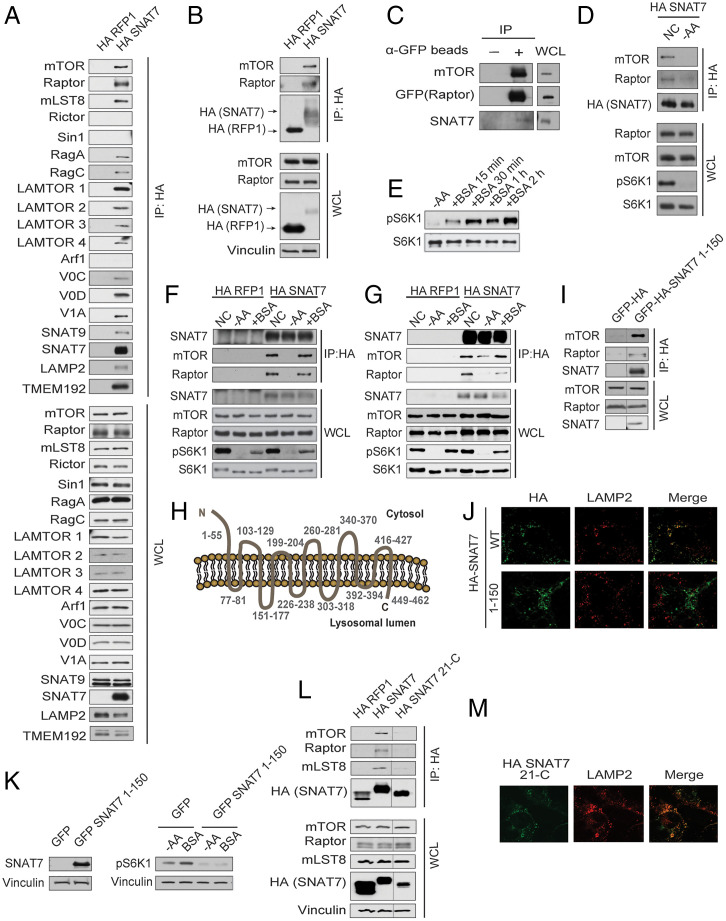
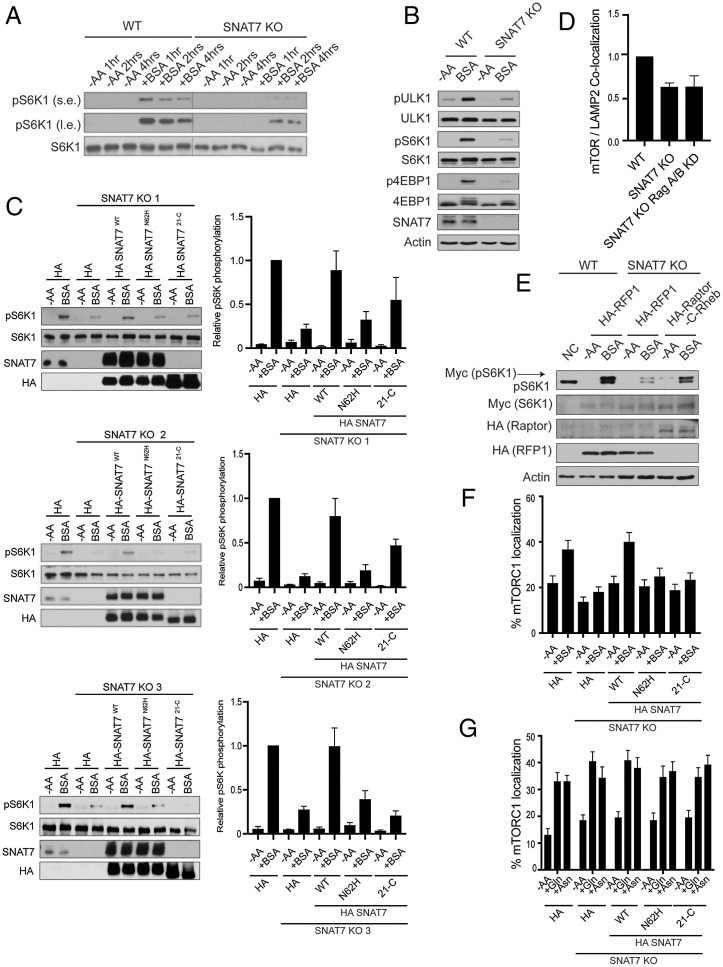
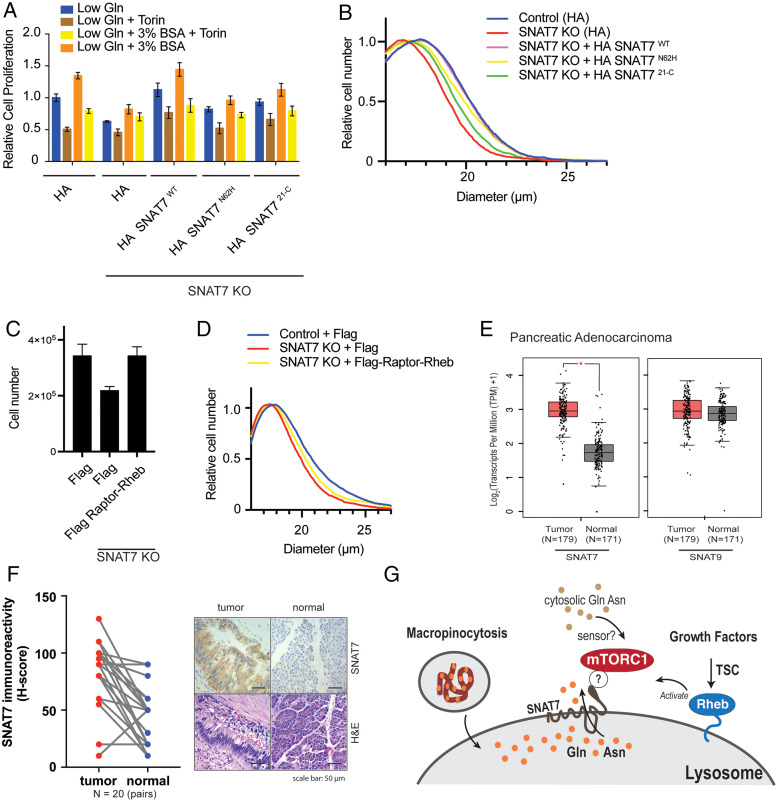
Mammalian target of rapamycin complex 1 (mTORC1) senses amino acids to control cell growth, metabolism, and autophagy. Some amino acids signal to mTORC1 through the Rag GTPase, whereas glutamine and asparagine activate mTORC1 through a Rag GTPase-independent pathway. Here, we show that the lysosomal glutamine and asparagine transporter SNAT7 activates mTORC1 after extracellular protein, such as albumin, is macropinocytosed. The N terminus of SNAT7 forms nutrient-sensitive interaction with mTORC1 and regulates mTORC1 activation independently of the Rag GTPases. Depletion of SNAT7 inhibits albumin-induced mTORC1 lysosomal localization and subsequent activation. Moreover, SNAT7 is essential to sustain KRAS-driven pancreatic cancer cell growth through mTORC1. Thus, SNAT7 links glutamine and asparagine signaling from extracellular protein to mTORC1 independently of the Rag GTPases and is required for macropinocytosis-mediated mTORC1 activation and pancreatic cancer cell growth.
Keywords: SNAT7; mTOR; macropinocytosis.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Saxton R. A., Sabatini D. M., mTOR signaling in growth, metabolism, and disease. Cell 169, 361–371 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
