

MYC drives aggressive prostate cancer by disrupting transcriptional pause release at androgen receptor targets
- PMID: 35562350
- PMCID: PMC9106722
- DOI: 10.1038/s41467-022-30257-z
MYC drives aggressive prostate cancer by disrupting transcriptional pause release at androgen receptor targets
Abstract
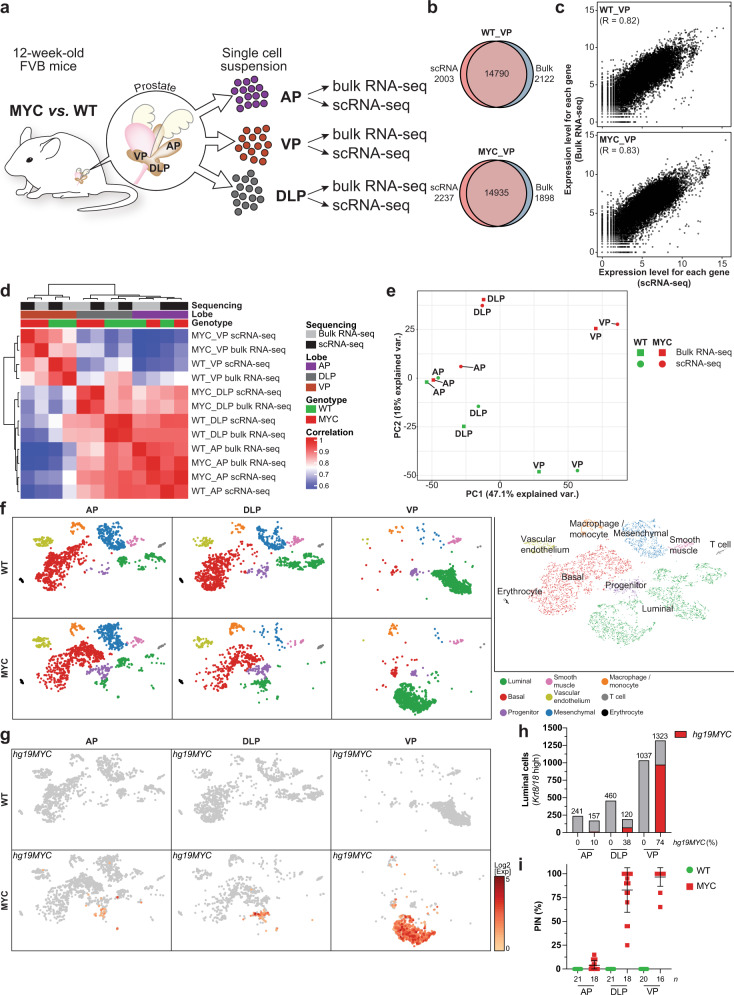
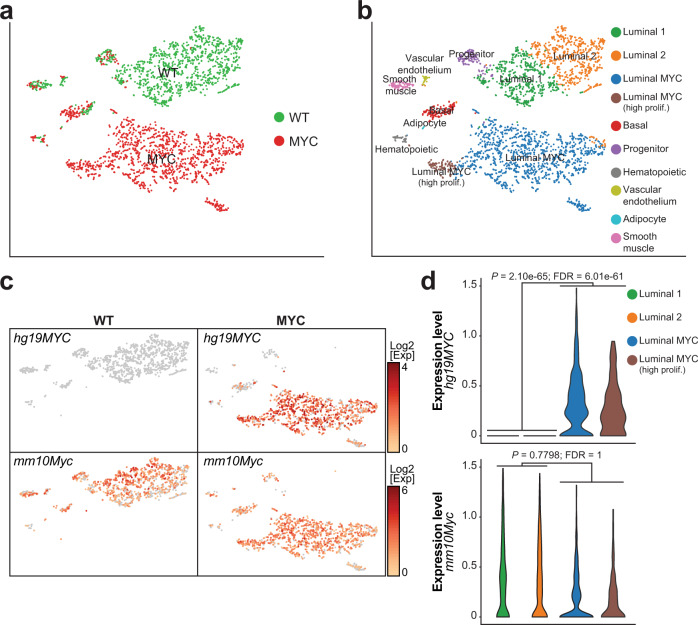
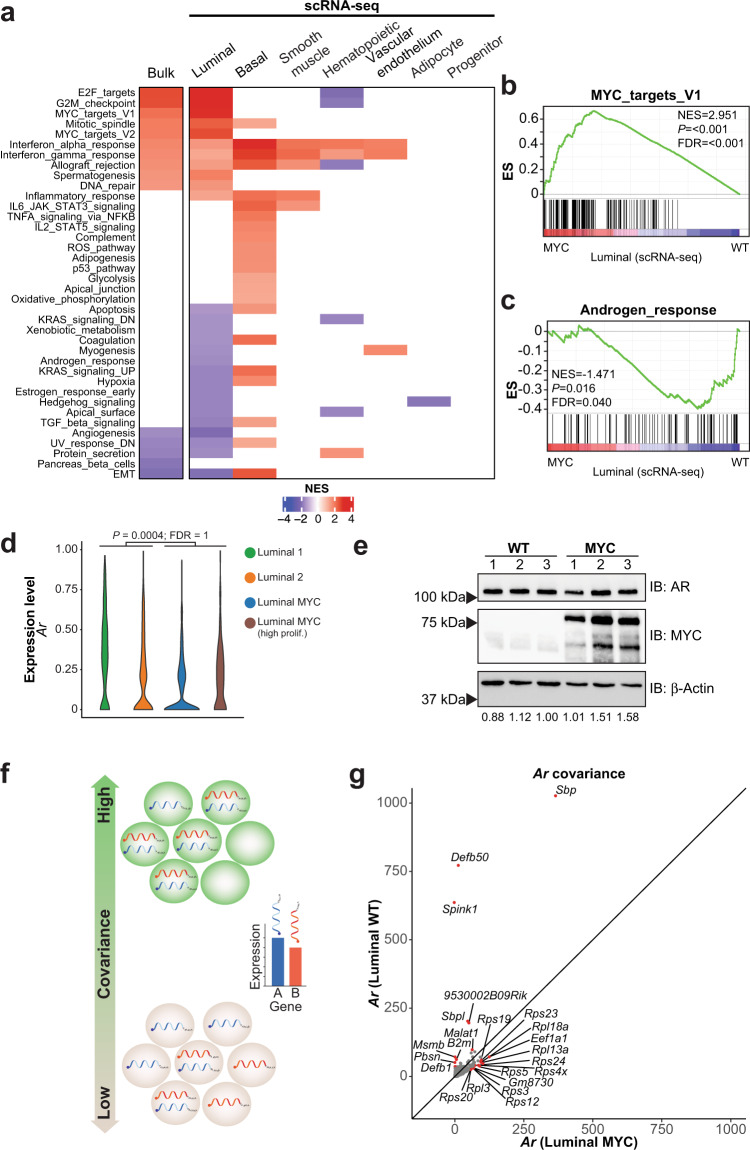
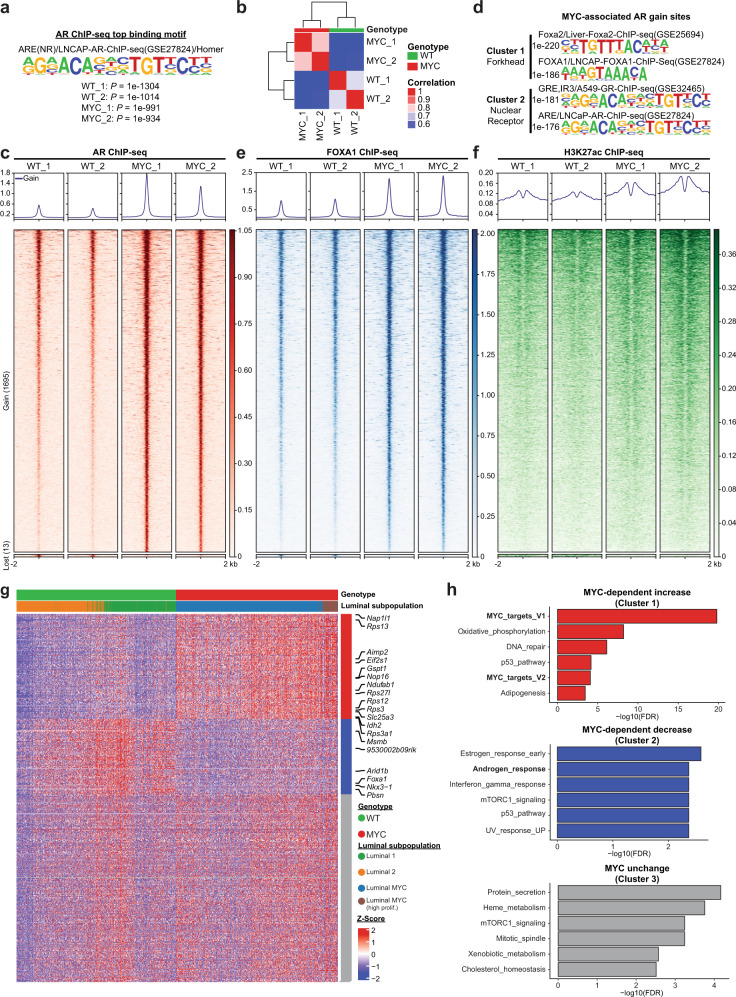
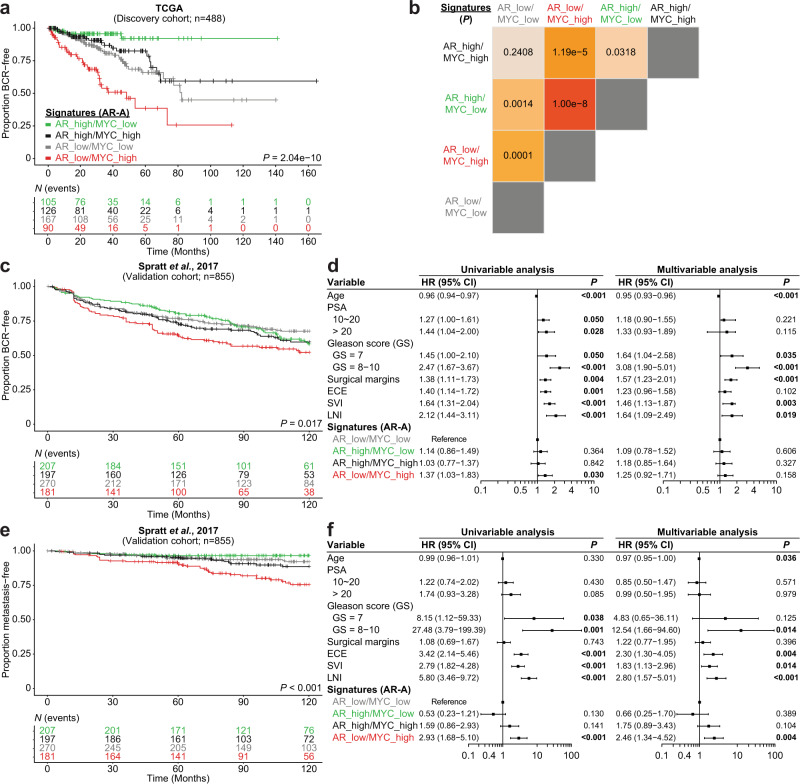
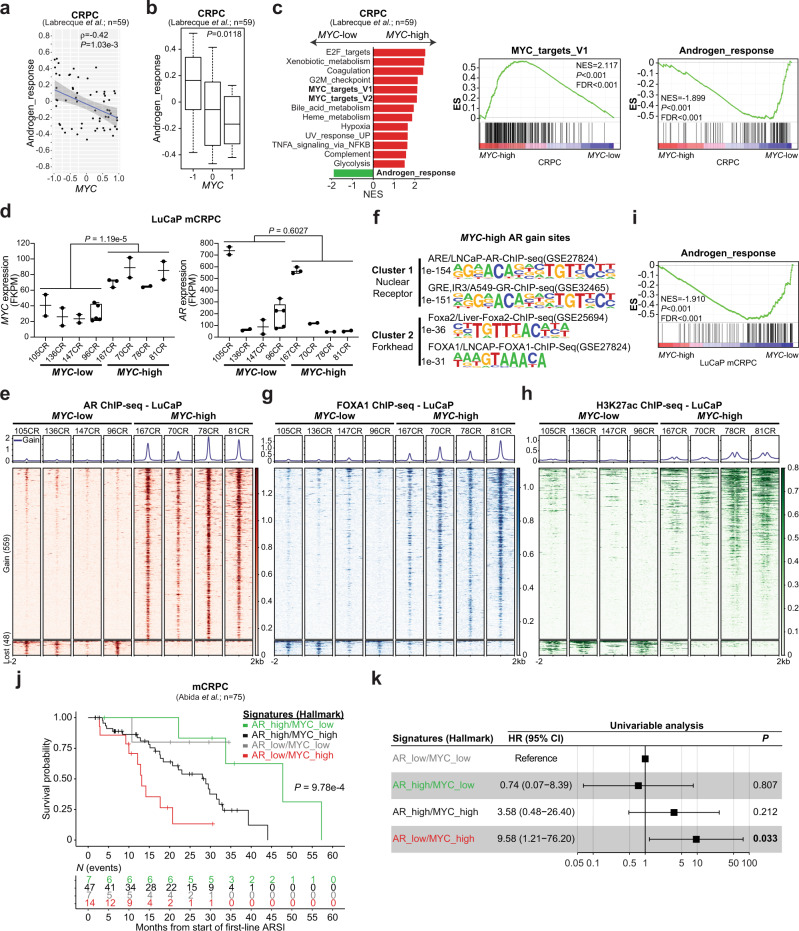
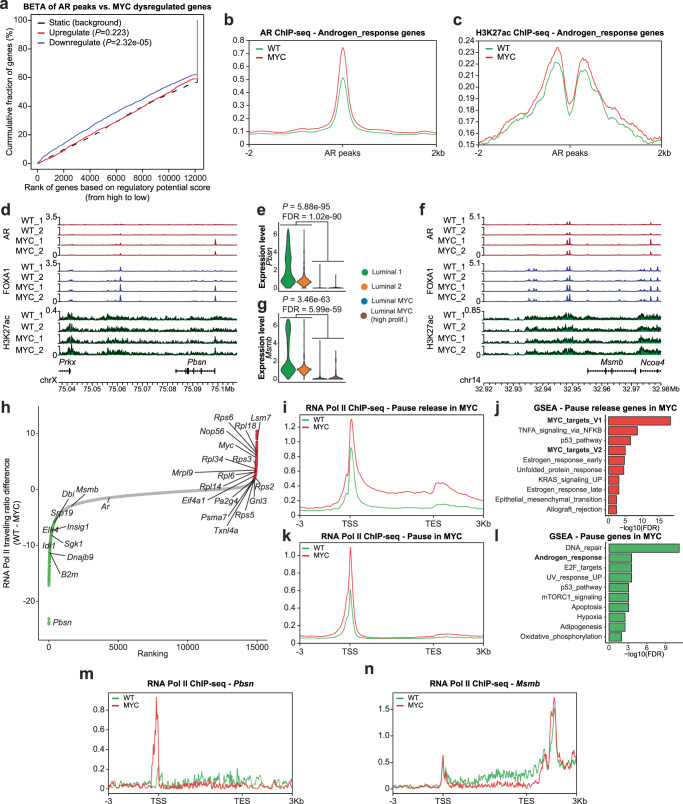
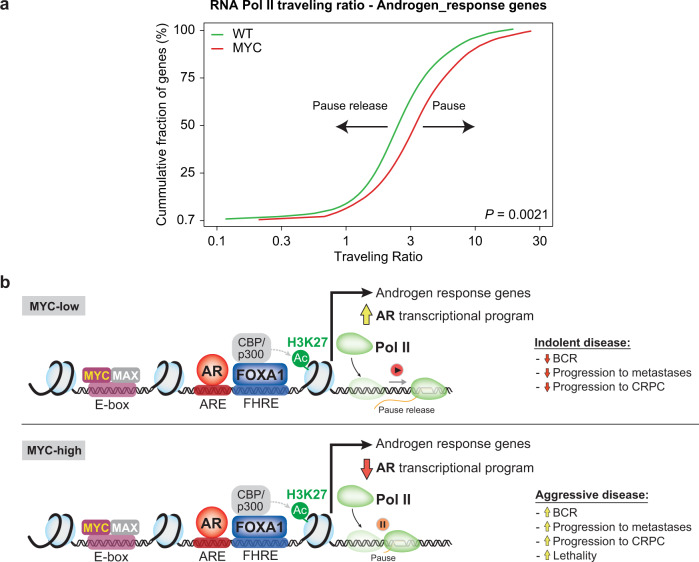
c-MYC (MYC) is a major driver of prostate cancer tumorigenesis and progression. Although MYC is overexpressed in both early and metastatic disease and associated with poor survival, its impact on prostate transcriptional reprogramming remains elusive. We demonstrate that MYC overexpression significantly diminishes the androgen receptor (AR) transcriptional program (the set of genes directly targeted by the AR protein) in luminal prostate cells without altering AR expression. Analyses of clinical specimens reveal that concurrent low AR and high MYC transcriptional programs accelerate prostate cancer progression toward a metastatic, castration-resistant disease. Data integration of single-cell transcriptomics together with ChIP-seq uncover an increase in RNA polymerase II (Pol II) promoter-proximal pausing at AR-dependent genes following MYC overexpression without an accompanying deactivation of AR-bound enhancers. Altogether, our findings suggest that MYC overexpression antagonizes the canonical AR transcriptional program and contributes to prostate tumor initiation and progression by disrupting transcriptional pause release at AR-regulated genes.
© 2022. The Author(s).
Conflict of interest statement
R.J.K. receive royalties from GenomeDx (now Veracyte) for Decipher testing. S.W. receives research funding from PreludeDX. K.W.W. serves on the scientific advisory board of T-Scan Therapeutics, SQZ Biotech, Nextechinvest and receives sponsored research funding from Novartis. He is a co-founder of Immunitas, a biotech company. These activities are not related to the research reported in this publication. D.E.S. receives personal fees from Janssen, AstraZeneca, and Blue Earth and funding from Janssen. E.C. received research funding under institutional SRA from Janssen Research and Development, Bayer Pharmaceuticals, KronosBio, Forma Pharmaceutics, Foghorn, Gilead, Sanofi, AbbVie, MacroGenics, and GSK. M.L.F. reports other support from Nuscan Diagnostics outside the submitted work. X.S.L. conducted the work while being a faculty at the Dana-Farber Cancer Institute and is currently a board member and CEO of GV20. M.B. and H.W.L. receives sponsored research support from Novartis. M.B. is a consultant to Aleta Biotherapeutics and H3 Biomedicine and serves on the SAB of Kronos Bio. The remaining authors declare no competing interests.
Figures
References
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J. Clin. 2019;69:7–34. - PubMed
-
- Huggins C, Hodges CV. Studies on prostatic cancer - I The effect of castration, of estrogen and of androgen injection on serum phosphatases in metastatic carcinoma of the prostate. Cancer Res. 1941;1:293–297. - PubMed
-
- Huggins C, Stevens RE, Hodges CV. Studies on prostate cancer II The effects of castration on advanced carcinoma of the prostate gland. Arch. Surg.-Chic. 1941;43:209–223. doi: 10.1001/archsurg.1941.01210140043004. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
