

Probing and Manipulating the Lateral Pressure Profile in Lipid Bilayers Using Membrane-Active Peptides-A Solid-State 19F NMR Study
- PMID: 35562938
- PMCID: PMC9101910
- DOI: 10.3390/ijms23094544
Probing and Manipulating the Lateral Pressure Profile in Lipid Bilayers Using Membrane-Active Peptides-A Solid-State 19F NMR Study
Abstract
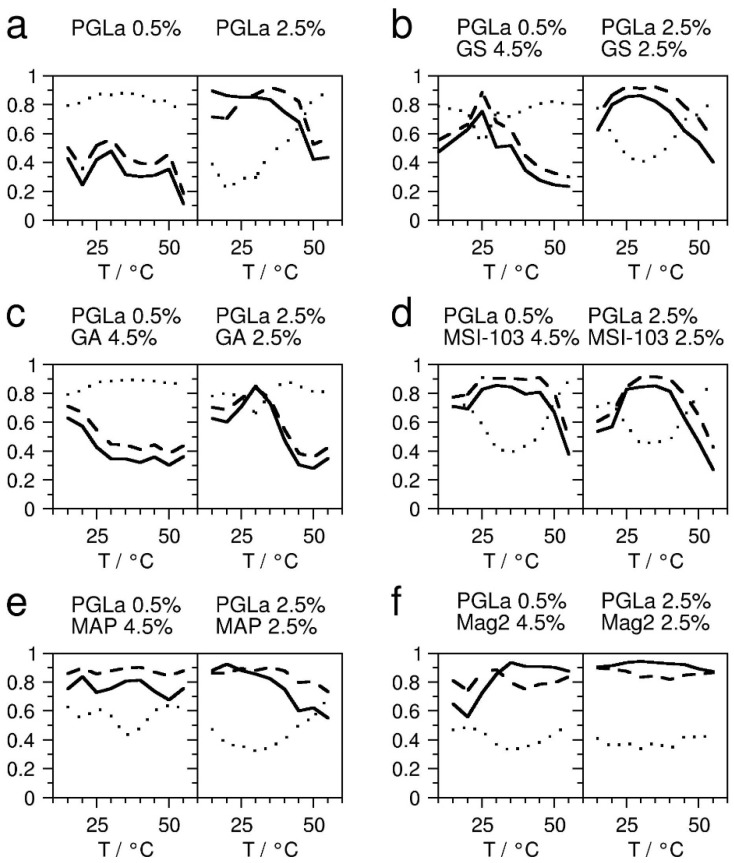
The lateral pressure profile constitutes an important physical property of lipid bilayers, influencing the binding, insertion, and function of membrane-active peptides, such as antimicrobial peptides. In this study, we demonstrate that the lateral pressure profile can be manipulated using the peptides residing in different regions of the bilayer. A 19F-labeled analogue of the amphiphilic peptide PGLa was used to probe the lateral pressure at different depths in the membrane. To evaluate the lateral pressure profile, we measured the orientation of this helical peptide with respect to the membrane using solid-state 19F-NMR, which is indicative of its degree of insertion into the bilayer. Using this experimental approach, we observed that the depth of insertion of the probe peptide changed in the presence of additional peptides and, furthermore, correlated with their location in the membrane. In this way, we obtained a tool to manipulate, as well as to probe, the lateral pressure profile in membranes.
Keywords: lateral pressure profile; lipid bilayer; membrane protein; membrane-active amphiphilic peptide; peptide crowding; solid-state 19F nuclear magnetic resonance.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures





Similar articles
-
Solid-state nuclear magnetic resonance relaxation studies of the interaction mechanism of antimicrobial peptides with phospholipid bilayer membranes.Biochemistry. 2005 Aug 2;44(30):10208-17. doi: 10.1021/bi050730p. Biochemistry. 2005. PMID: 16042398
-
Solid-state NMR investigation of the depth of insertion of protegrin-1 in lipid bilayers using paramagnetic Mn2+.Biophys J. 2003 Oct;85(4):2363-73. doi: 10.1016/S0006-3495(03)74660-8. Biophys J. 2003. PMID: 14507700 Free PMC article.
-
Diphytanoyl lipids as model systems for studying membrane-active peptides.Biochim Biophys Acta Biomembr. 2017 Oct;1859(10):1828-1837. doi: 10.1016/j.bbamem.2017.06.003. Epub 2017 Jun 3. Biochim Biophys Acta Biomembr. 2017. PMID: 28587828
-
Elastic deformation and area per lipid of membranes: atomistic view from solid-state deuterium NMR spectroscopy.Biochim Biophys Acta. 2015 Jan;1848(1 Pt B):246-59. doi: 10.1016/j.bbamem.2014.06.004. Epub 2014 Jun 16. Biochim Biophys Acta. 2015. PMID: 24946141 Free PMC article. Review.
-
Solid-state (19)F-NMR of peptides in native membranes.Top Curr Chem. 2012;306:89-118. doi: 10.1007/128_2011_162. Top Curr Chem. 2012. PMID: 21598096 Review.
Cited by
-
Membranolytic Mechanism of Amphiphilic Antimicrobial β-Stranded [KL]n Peptides.Biomedicines. 2022 Aug 24;10(9):2071. doi: 10.3390/biomedicines10092071. Biomedicines. 2022. PMID: 36140173 Free PMC article.
-
Dynamic Structures of Bioactive Proteins as Determined by Nuclear Magnetic Resonance.Int J Mol Sci. 2023 Dec 25;25(1):295. doi: 10.3390/ijms25010295. Int J Mol Sci. 2023. PMID: 38203465 Free PMC article.
-
Investigating How Lysophosphatidylcholine and Lysophosphatidylethanolamine Enhance the Membrane Permeabilization Efficacy of Host Defense Peptide Piscidin 1.J Phys Chem B. 2025 Jan 9;129(1):210-227. doi: 10.1021/acs.jpcb.4c05845. Epub 2024 Dec 16. J Phys Chem B. 2025. PMID: 39681296
References
-
- Cantor R.S. Lateral pressures in cell membranes: A mechanism for modulation of protein function. J. Phys. Chem. B. 1997;101:1723–1725. doi: 10.1021/jp963911x. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
