

Overexpression of EgrIAA20 from Eucalyptus grandis, a Non-Canonical Aux/ IAA Gene, Specifically Decouples Lignification of the Different Cell-Types in Arabidopsis Secondary Xylem
- PMID: 35563457
- PMCID: PMC9100763
- DOI: 10.3390/ijms23095068
Overexpression of EgrIAA20 from Eucalyptus grandis, a Non-Canonical Aux/ IAA Gene, Specifically Decouples Lignification of the Different Cell-Types in Arabidopsis Secondary Xylem
Abstract
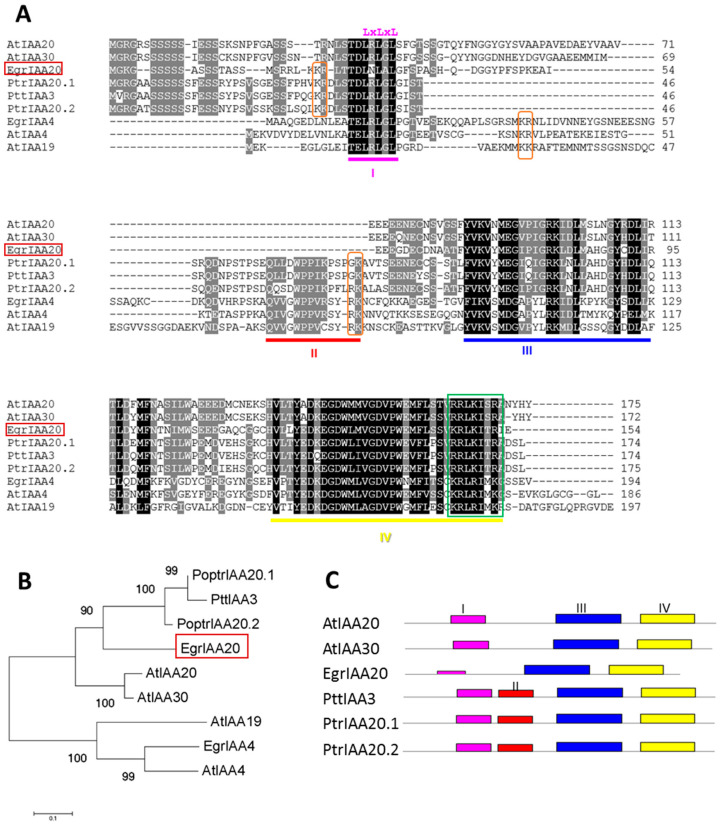
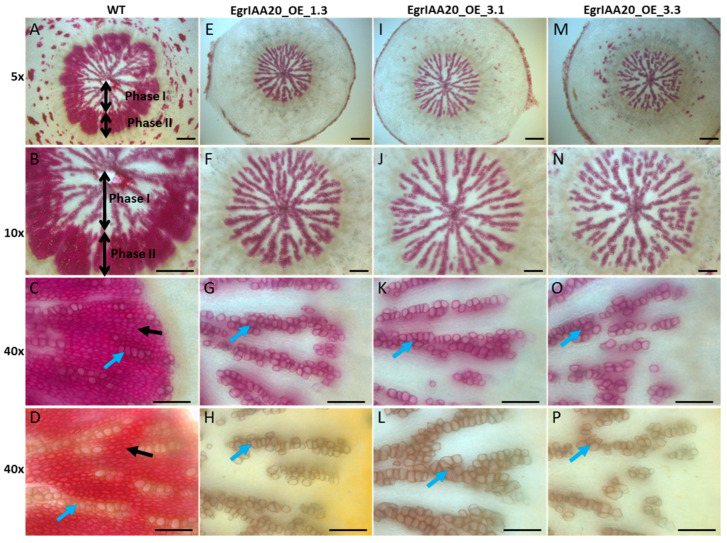
Wood (secondary xylem) formation is regulated by auxin, which plays a pivotal role as an integrator of developmental and environmental cues. However, our current knowledge of auxin-signaling during wood formation is incomplete. Our previous genome-wide analysis of Aux/IAAs in Eucalyptus grandis showed the presence of the non-canonical paralog member EgrIAA20 that is preferentially expressed in cambium. We analyzed its cellular localization using a GFP fusion protein and its transcriptional activity using transactivation assays, and demonstrated its nuclear localization and strong auxin response repressor activity. In addition, we functionally tested the role of EgrIAA20 by constitutive overexpression in Arabidopsis to investigate for phenotypic changes in secondary xylem formation. Transgenic Arabidopsis plants overexpressing EgrIAA20 were smaller and displayed impaired development of secondary fibers, but not of other wood cell types. The inhibition in fiber development specifically affected their cell wall lignification. We performed yeast-two-hybrid assays to identify EgrIAA20 protein partners during wood formation in Eucalyptus, and identified EgrIAA9A, whose ortholog PtoIAA9 in poplar is also known to be involved in wood formation. Altogether, we showed that EgrIAA20 is an important auxin signaling component specifically involved in controlling the lignification of wood fibers.
Keywords: Arabidopsis; Eucalyptus; IAA20; auxin; cambium differentiation; non-canonical Aux/IAA; secondary fiber; secondary xylem; syringyl lignin; wood.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures







Similar articles
-
Comprehensive genome-wide analysis of the Aux/IAA gene family in Eucalyptus: evidence for the role of EgrIAA4 in wood formation.Plant Cell Physiol. 2015 Apr;56(4):700-14. doi: 10.1093/pcp/pcu215. Epub 2015 Jan 9. Plant Cell Physiol. 2015. PMID: 25577568
-
Eucalyptus grandis AUX/INDOLE-3-ACETIC ACID 13 (EgrIAA13) is a novel transcriptional regulator of xylogenesis.Plant Mol Biol. 2022 May;109(1-2):51-65. doi: 10.1007/s11103-022-01255-y. Epub 2022 Mar 16. Plant Mol Biol. 2022. PMID: 35292886 Free PMC article.
-
Auxin-mediated Aux/IAA-ARF-HB signaling cascade regulates secondary xylem development in Populus.New Phytol. 2019 Apr;222(2):752-767. doi: 10.1111/nph.15658. Epub 2019 Feb 1. New Phytol. 2019. PMID: 30582614
-
Current Understanding of the Genetics and Molecular Mechanisms Regulating Wood Formation in Plants.Genes (Basel). 2022 Jun 30;13(7):1181. doi: 10.3390/genes13071181. Genes (Basel). 2022. PMID: 35885964 Free PMC article. Review.
-
Arabidopsis as a model for wood formation.Curr Opin Biotechnol. 2011 Apr;22(2):293-9. doi: 10.1016/j.copbio.2010.11.008. Epub 2010 Dec 6. Curr Opin Biotechnol. 2011. PMID: 21144727 Review.
Cited by
-
Transcriptomic Insights into Functions of LkABCG36 and LkABCG40 in Nicotiana tabacum.Plants (Basel). 2023 Jan 4;12(2):227. doi: 10.3390/plants12020227. Plants (Basel). 2023. PMID: 36678941 Free PMC article.
-
A non-canonical Aux/IAA gene MsIAA32 regulates peltate glandular trichome development in spearmint.Front Plant Sci. 2024 Feb 5;15:1284125. doi: 10.3389/fpls.2024.1284125. eCollection 2024. Front Plant Sci. 2024. PMID: 38375083 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
