

Epigenetic regulation of innate immune memory in microglia
- PMID: 35568856
- PMCID: PMC9107649
- DOI: 10.1186/s12974-022-02463-5
Epigenetic regulation of innate immune memory in microglia
Abstract
Background: Microglia are the tissue-resident macrophages of the CNS. They originate in the yolk sac, colonize the CNS during embryonic development and form a self-sustaining population with limited turnover. A consequence of their relative slow turnover is that microglia can serve as a long-term memory for inflammatory or neurodegenerative events.
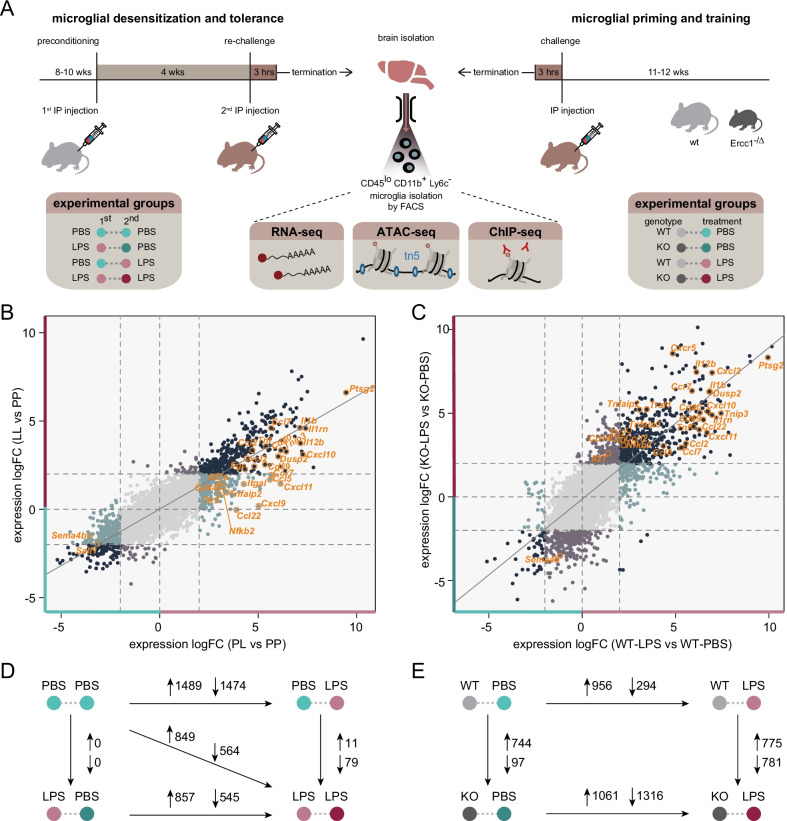
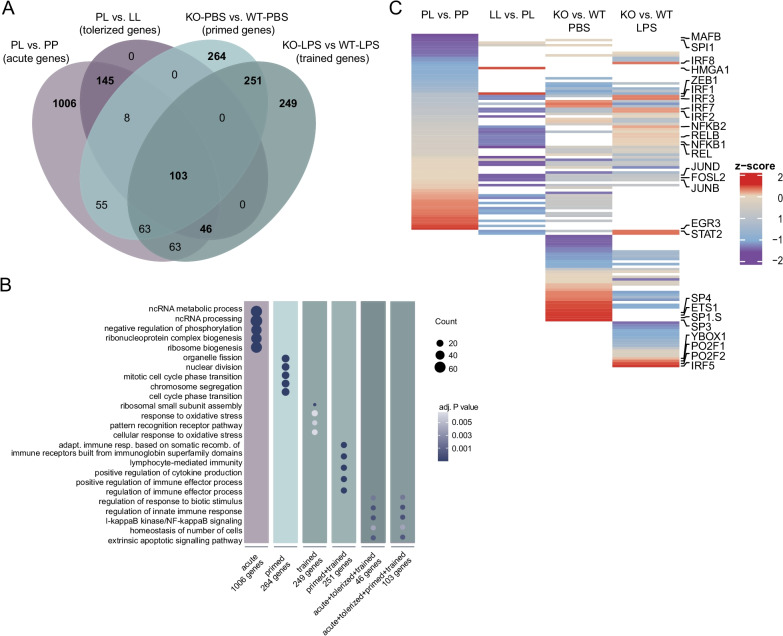
Methods: Using ATAC-, ChIP- and RNA-sequencing, we characterized the epigenomes and transcriptomes of FACS-purified microglia from mice exposed to different stimuli. A repeated endotoxin challenge (LPS) was used to induce tolerance in microglia, while genotoxic stress (DNA repair deficiency-induced accelerated aging through Ercc1 deficiency) resulted in primed (hypersensitive) microglia.
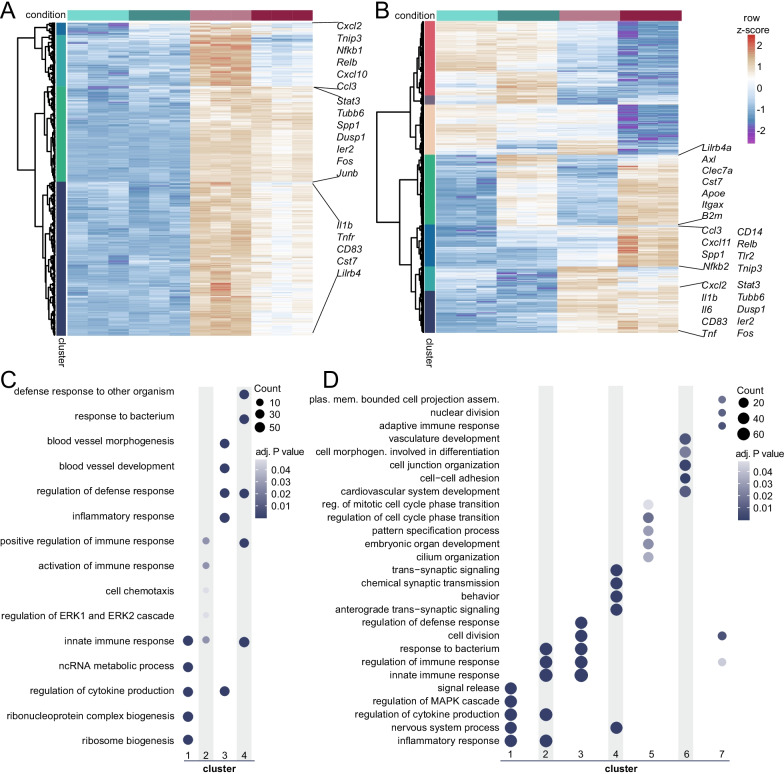
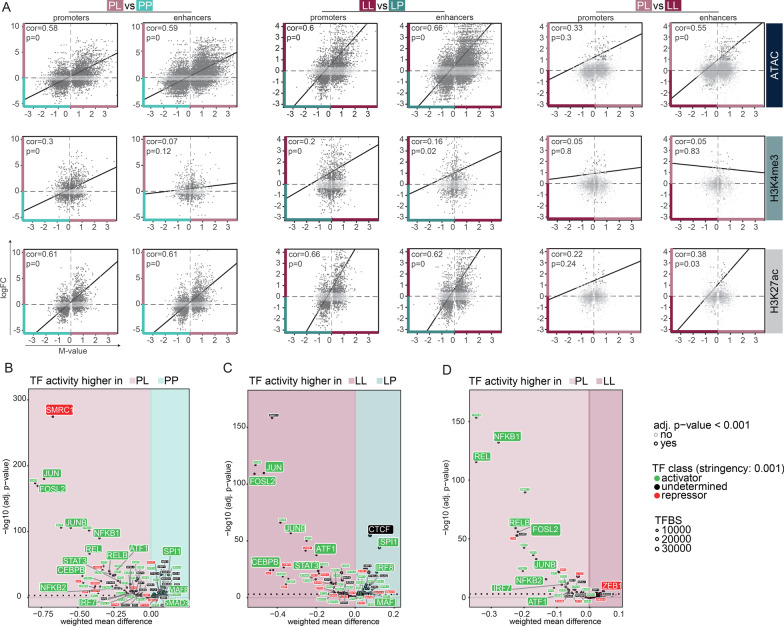
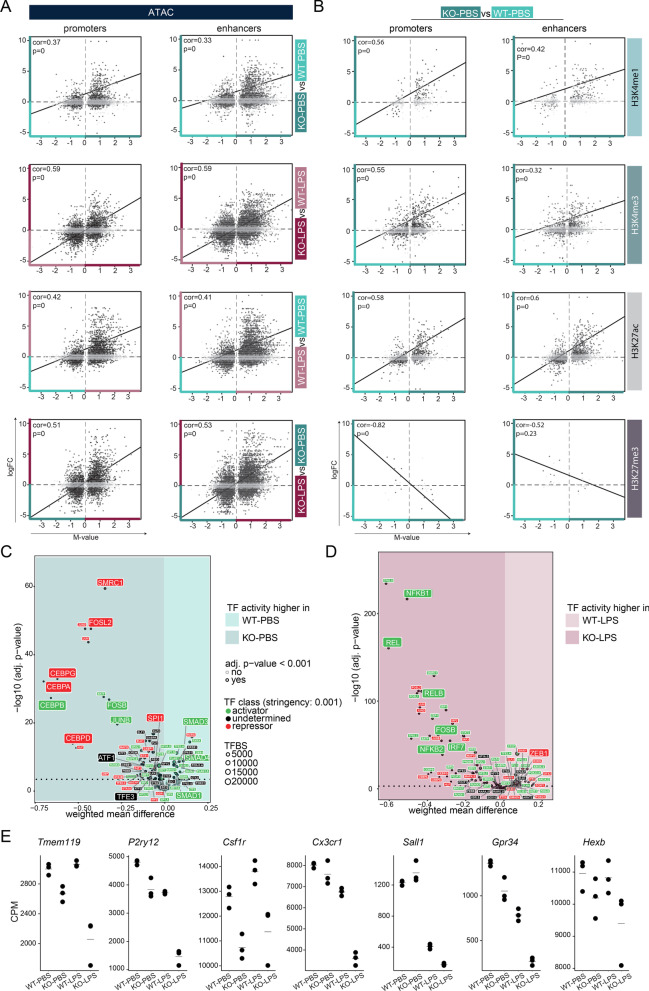
Results: Whereas the enrichment of permissive epigenetic marks at enhancer regions could explain training (hyper-responsiveness) of primed microglia to an LPS challenge, the tolerized response of microglia seems to be regulated by loss of permissive epigenetic marks. We identify that inflammatory stimuli and accelerated aging as a result of genotoxic stress activate distinct gene networks. These gene networks and associated biological processes are partially overlapping, which is likely driven by specific transcription factor networks, resulting in altered epigenetic signatures and distinct functional (desensitized vs. primed) microglia phenotypes.
Conclusion: This study provides insight into epigenetic profiles and transcription factor networks associated with transcriptional signatures of tolerized and trained microglia in vivo, leading to a better understanding of innate immune memory of microglia.
Keywords: Chromatin; Innate immunity; Microglia; Neuroinflammation; Priming; Tolerance.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
