

Complex Ecotype Dynamics Evolve in Response to Fluctuating Resources
- PMID: 35575545
- PMCID: PMC9239185
- DOI: 10.1128/mbio.03467-21
Complex Ecotype Dynamics Evolve in Response to Fluctuating Resources
Abstract
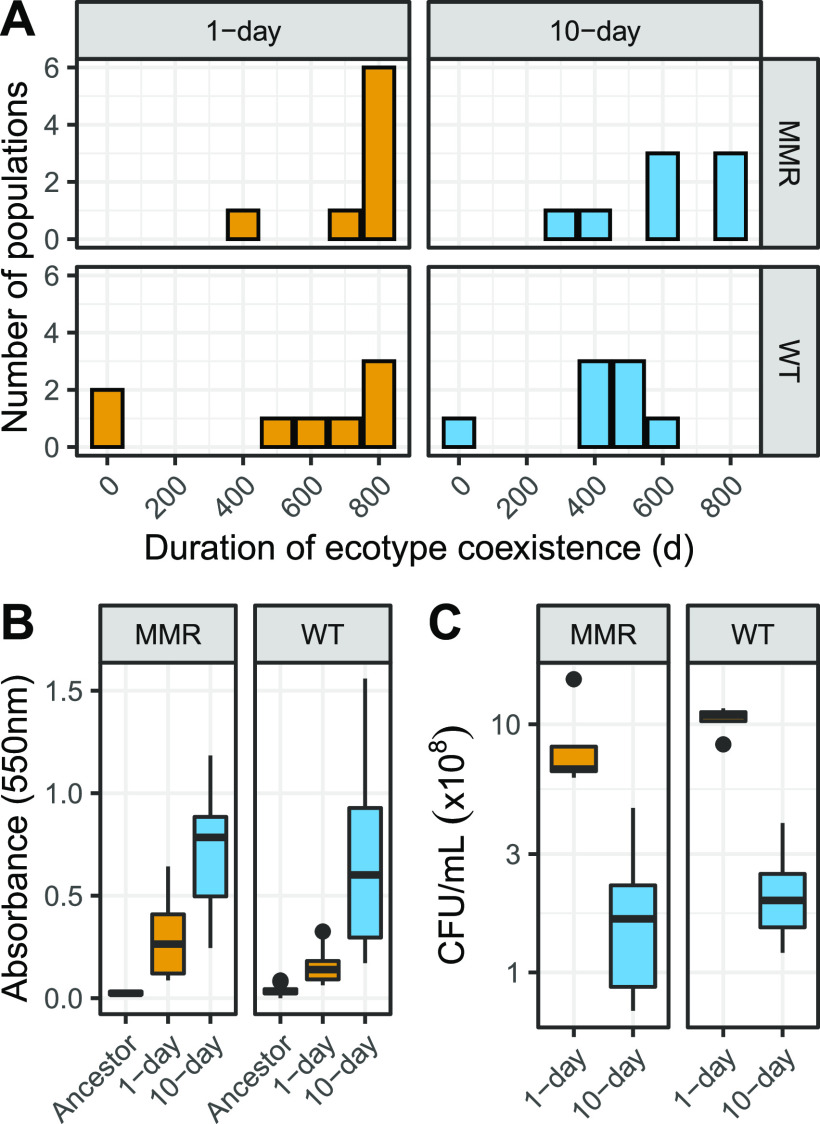
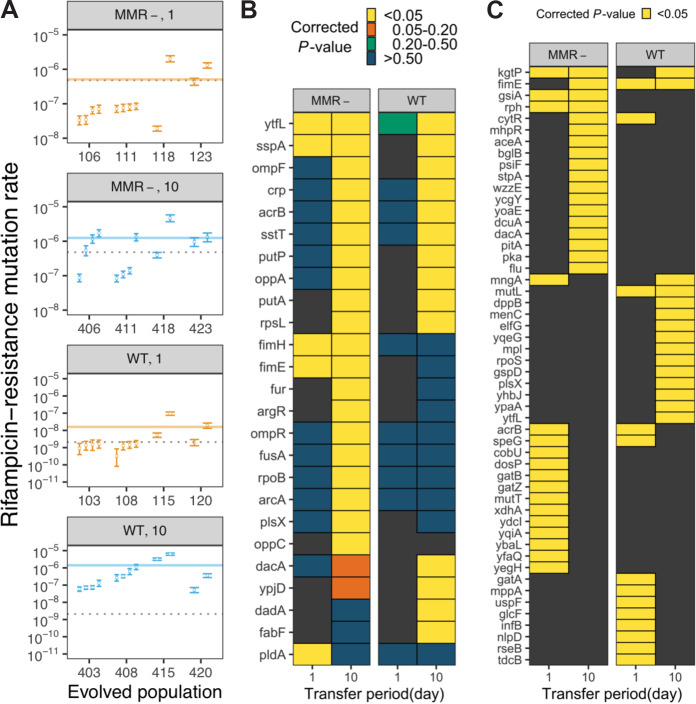
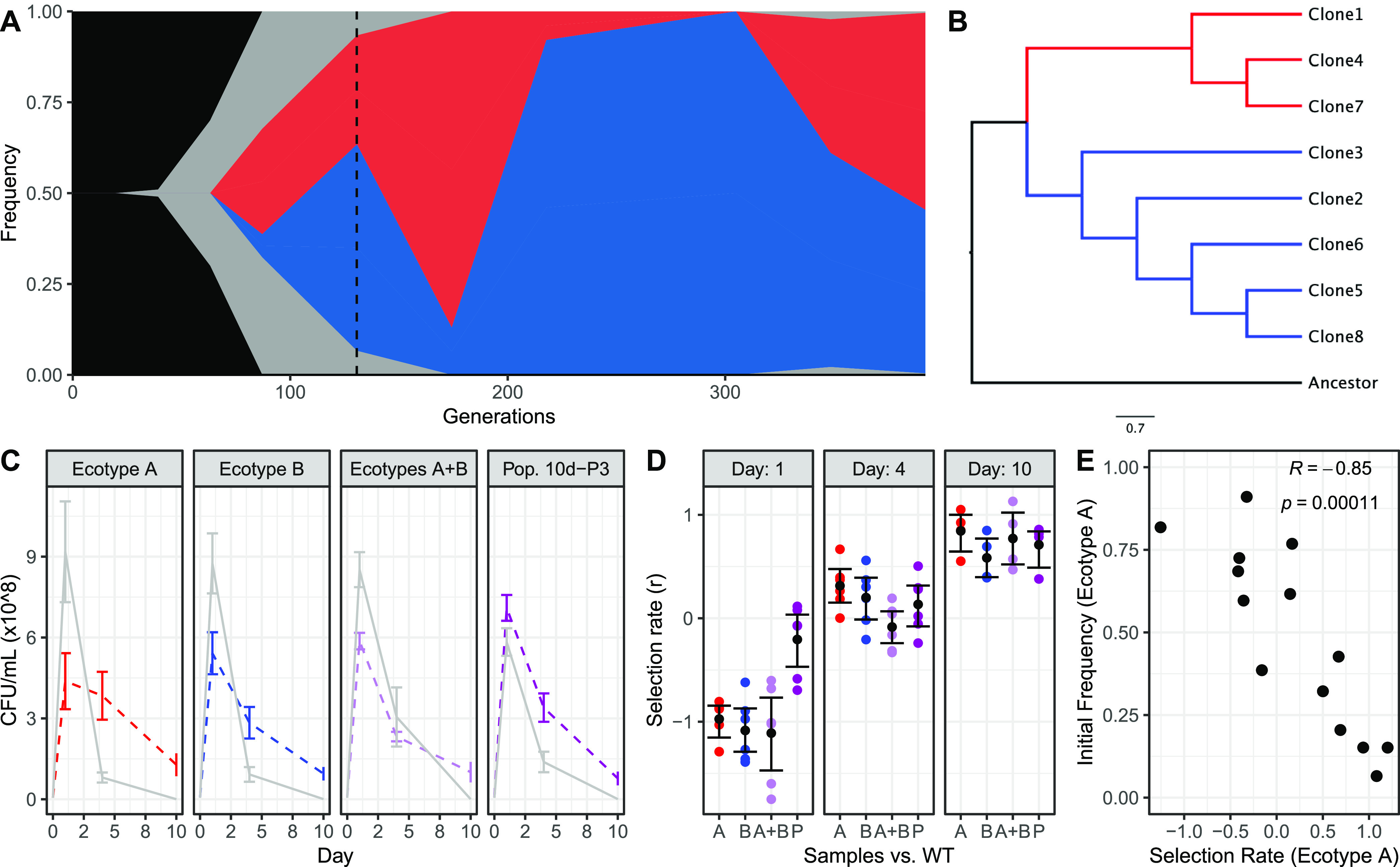
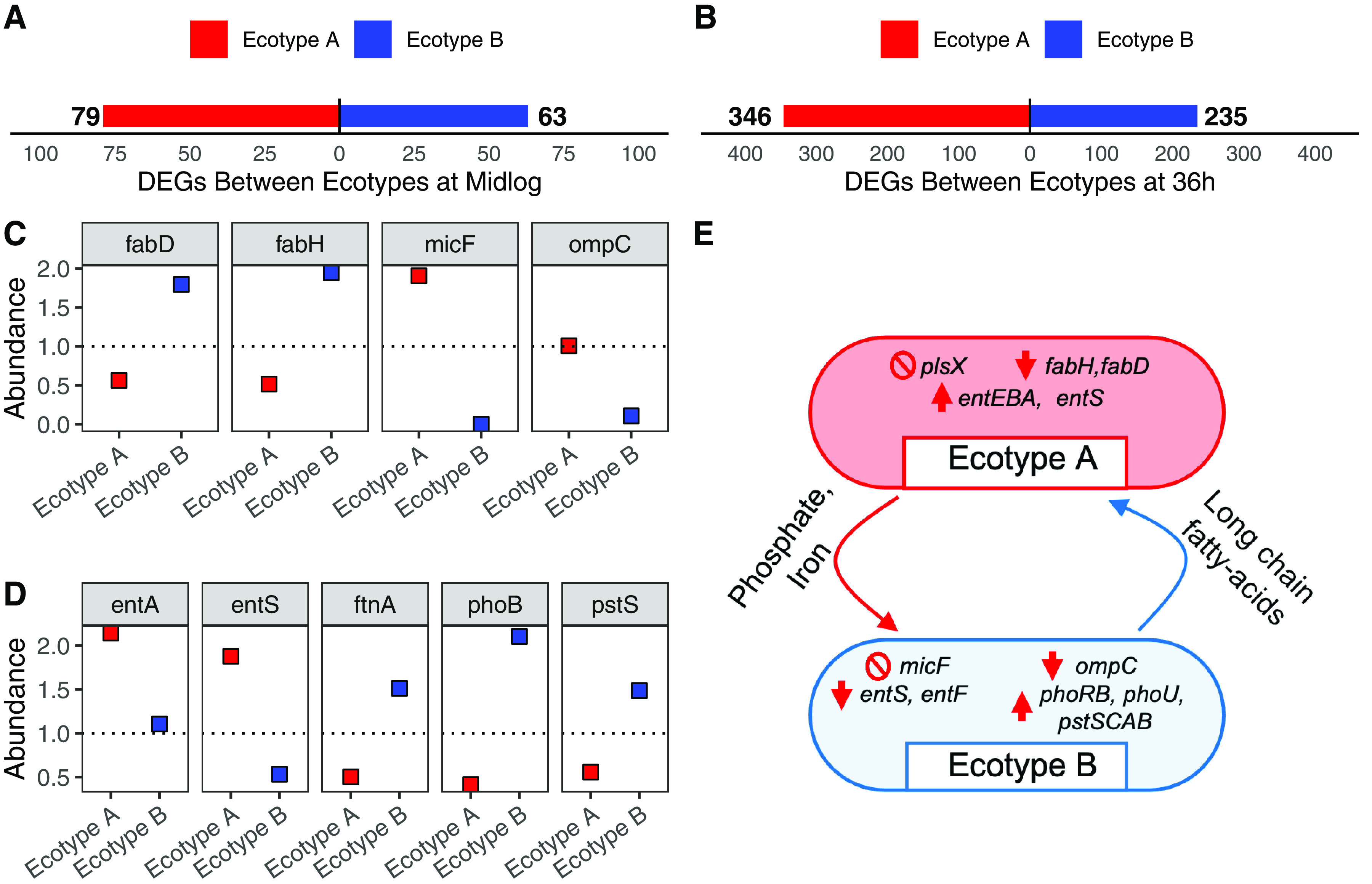
Ecotypic diversification and its associated cooperative behaviors are frequently observed in natural microbial populations whose access to resources is often sporadic. However, the extent to which fluctuations in resource availability influence the emergence of cooperative ecotypes is not fully understood. To determine how exposure to repeated resource limitation affects the establishment and long-term maintenance of ecotypes in a structured environment, we followed 32 populations of Escherichia coli evolving to either 1-day or 10-day feast/famine cycles for 900 days. Population-level analysis revealed that compared to populations evolving to 1-day cycles, 10-day populations evolved increased biofilm density, higher parallelism in mutational targets, and increased mutation rates. As previous investigations of evolution in structured environments have identified biofilm formation as the earliest observable phenotype associated with diversification of ecotypes, we revived cultures midway through the evolutionary process and conducted additional genomic, transcriptional, and phenotypic analyses of clones isolated from these evolving populations. We found not only that 10-day feast/famine cycles support multiple ecotypes but also that these ecotypes exhibit cooperative behavior. Consistent with the black queen hypothesis, or evolution of cooperation by gene loss, transcriptomic evidence suggests the evolution of bidirectional cross-feeding behaviors based on essential resources. These results provide insight into how analogous cooperative relationships may emerge in natural microbial communities. IMPORTANCE Despite regular feast and famine conditions representing an environmental pressure that is commonly encountered by microbial communities, the evolutionary outcomes of repeated cycles of feast and famine have been less studied. By experimentally evolving initially isogenic Escherichia coli populations to 10-day feast/famine cycles, we observed rapid diversification into ecotypes with evidence of bidirectional cross-feeding on costly resources and frequency-dependent fitness. Although unidirectional cross-feeding has been repeatedly observed to evolve in laboratory culture, most investigations of bidirectional cooperative behaviors in microbial populations have been conducted in engineered communities. This work demonstrates the de novo evolution of black queen relationships in a microbial population originating from a single ancestor, providing a model for investigation of the eco-evolutionary processes leading to mutualistic cooperation.
Keywords: Escherichia coli; black queen hypothesis; experimental evolution; intraspecific cooperation; starvation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
