

A Silent Operon of Photorhabdus luminescens Encodes a Prodrug Mimic of GTP
- PMID: 35575547
- PMCID: PMC9239236
- DOI: 10.1128/mbio.00700-22
A Silent Operon of Photorhabdus luminescens Encodes a Prodrug Mimic of GTP
Abstract
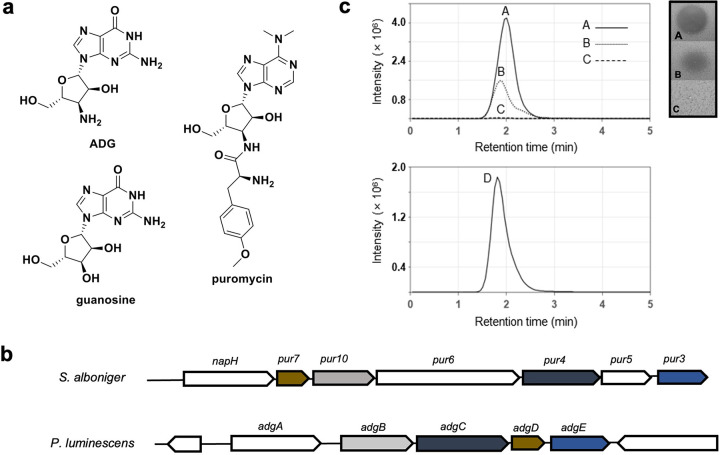
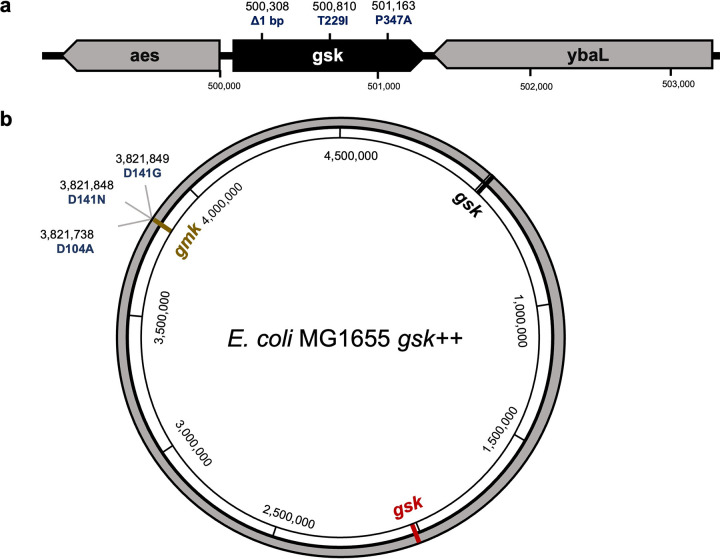
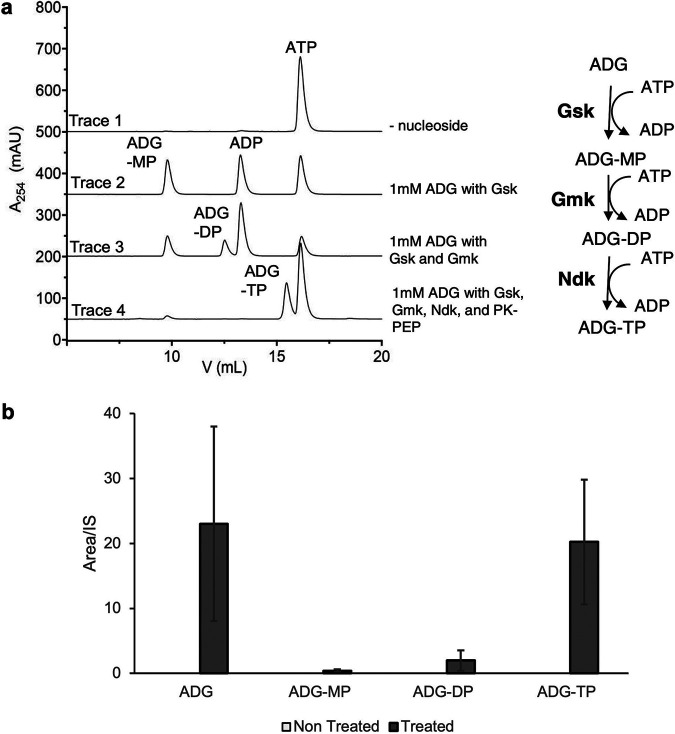
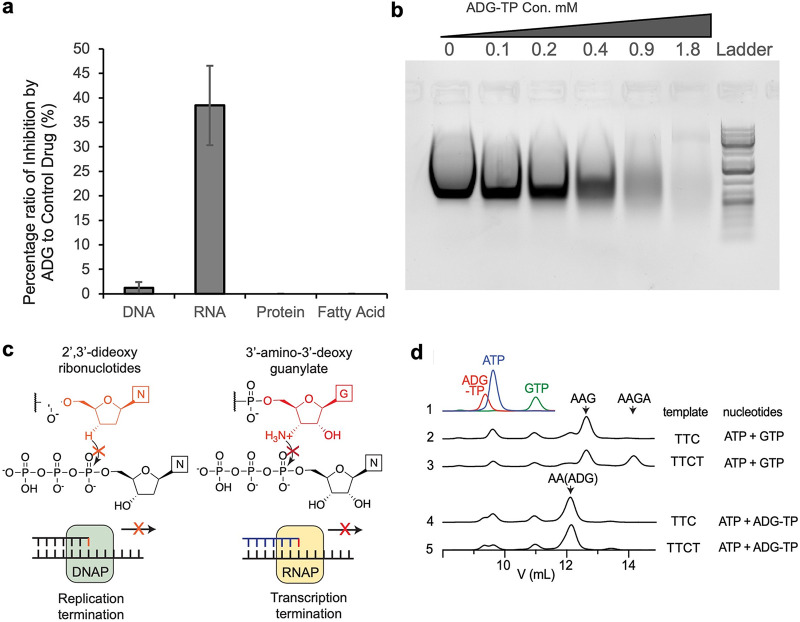
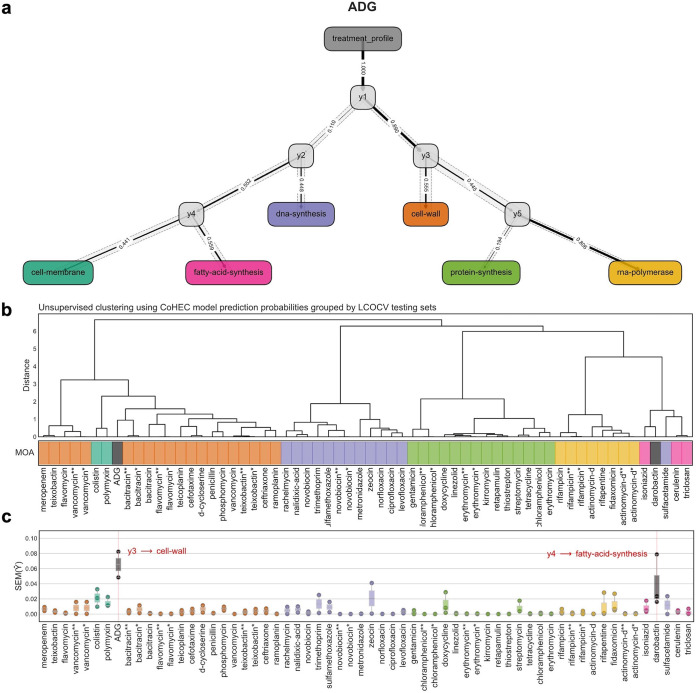
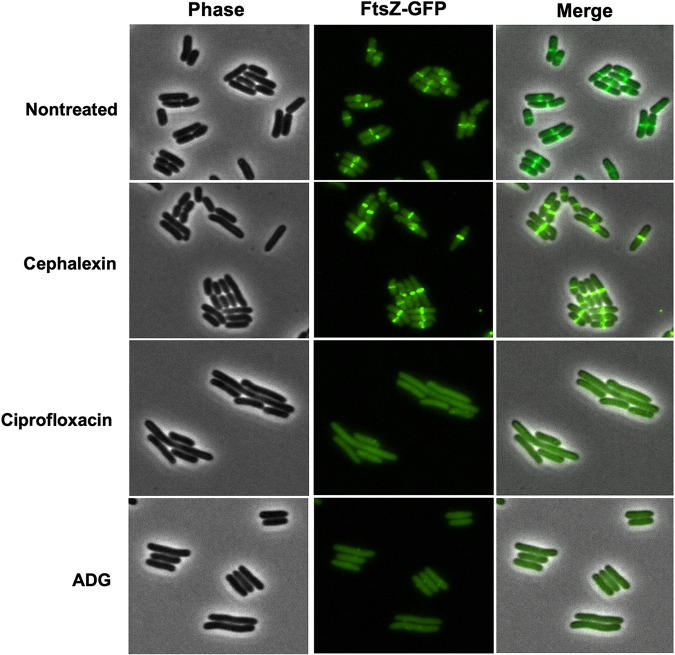
With the overmining of actinomycetes for compounds acting against Gram-negative pathogens, recent efforts to discover novel antibiotics have been focused on other groups of bacteria. Teixobactin, the first antibiotic without detectable resistance that binds lipid II, comes from an uncultured Eleftheria terra, a betaproteobacterium; odilorhabdins, from Xenorhabdus, are broad-spectrum inhibitors of protein synthesis, and darobactins from Photorhabdus target BamA, the essential chaperone of the outer membrane of Gram-negative bacteria. Xenorhabdus and Photorhabdus are symbionts of the nematode gut microbiome and attractive producers of secondary metabolites. Only small portions of their biosynthetic gene clusters (BGC) are expressed in vitro. To access their silent operons, we first separated extracts from a small library of isolates into fractions, resulting in 200-fold concentrated material, and then screened them for antimicrobial activity. This resulted in a hit with selective activity against Escherichia coli, which we identified as a novel natural product antibiotic, 3'-amino 3'-deoxyguanosine (ADG). Mutants resistant to ADG mapped to gsk and gmk, kinases of guanosine. Biochemical analysis shows that ADG is a prodrug that is converted into an active ADG triphosphate (ADG-TP), a mimic of GTP. ADG incorporates into a growing RNA chain, interrupting transcription, and inhibits cell division, apparently by interfering with the GTPase activity of FtsZ. Gsk of the purine salvage pathway, which is the first kinase in the sequential phosphorylation of ADG, is restricted to E. coli and closely related species, explaining the selectivity of the compound. There are probably numerous targets of ADG-TP among GTP-dependent proteins. The discovery of ADG expands our knowledge of prodrugs, which are rare among natural compounds. IMPORTANCE Drug-resistant Gram-negative bacteria have become the major problem driving the antimicrobial resistance crisis. Searching outside the overmined actinomycetes, we focused on Photorhabdus, gut symbionts of enthomopathogenic nematodes that carry up to 40 biosynthetic gene clusters coding for secondary metabolites. Most of these are silent and do not express in vitro. To gain access to silent operons, we first fractionated supernatant from Photorhabdus and then tested 200-fold concentrated material for activity. This resulted in the isolation of a novel antimicrobial, 3'-amino 3'-deoxyguanosine (ADG), active against E. coli. ADG is an analog of guanosine and is converted into an active ADG-TP in the cell. ADG-TP inhibits transcription and probably numerous other GTP-dependent targets, such as FtsZ. Natural product prodrugs have been uncommon; discovery of ADG broadens our knowledge of this type of antibiotic.
Keywords: antibiotic resistance; natural product; nucleoside analog.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Exploring Xenorhabdus and Photorhabdus Nematode Symbionts in Search of Novel Therapeutics.Molecules. 2024 Oct 31;29(21):5151. doi: 10.3390/molecules29215151. Molecules. 2024. PMID: 39519791 Free PMC article. Review.
-
A study on Xenorhabdus and Photorhabdus isolates from Northeastern Thailand: Identification, antibacterial activity, and association with entomopathogenic nematode hosts.PLoS One. 2021 Aug 12;16(8):e0255943. doi: 10.1371/journal.pone.0255943. eCollection 2021. PLoS One. 2021. PMID: 34383819 Free PMC article.
-
Small molecule produced by Photorhabdus interferes with ubiquinone biosynthesis in Gram-negative bacteria.mBio. 2024 Oct 16;15(10):e0116724. doi: 10.1128/mbio.01167-24. Epub 2024 Sep 10. mBio. 2024. PMID: 39254306 Free PMC article.
-
A new antibiotic selectively kills Gram-negative pathogens.Nature. 2019 Dec;576(7787):459-464. doi: 10.1038/s41586-019-1791-1. Epub 2019 Nov 20. Nature. 2019. PMID: 31747680 Free PMC article.
-
Refining the Natural Product Repertoire in Entomopathogenic Bacteria.Trends Microbiol. 2018 Oct;26(10):833-840. doi: 10.1016/j.tim.2018.04.007. Epub 2018 May 22. Trends Microbiol. 2018. PMID: 29801772 Review.
Cited by
-
Sophisticated natural products as antibiotics.Nature. 2024 Aug;632(8023):39-49. doi: 10.1038/s41586-024-07530-w. Epub 2024 Jul 31. Nature. 2024. PMID: 39085542 Free PMC article. Review.
-
Anti-Klebsiella pneumoniae activity of secondary metabolism of Achromobacter from the intestine of Periplaneta americana.BMC Microbiol. 2023 Jun 5;23(1):162. doi: 10.1186/s12866-023-02909-7. BMC Microbiol. 2023. PMID: 37277707 Free PMC article.
-
Escherichia coli has an undiscovered ability to inhibit the growth of both Gram-negative and Gram-positive bacteria.Sci Rep. 2024 Mar 28;14(1):7420. doi: 10.1038/s41598-024-57996-x. Sci Rep. 2024. PMID: 38548840 Free PMC article.
-
Exploring Xenorhabdus and Photorhabdus Nematode Symbionts in Search of Novel Therapeutics.Molecules. 2024 Oct 31;29(21):5151. doi: 10.3390/molecules29215151. Molecules. 2024. PMID: 39519791 Free PMC article. Review.
-
The Search for Antibacterial Inhibitors Targeting Cell Division Protein FtsZ at Its Nucleotide and Allosteric Binding Sites.Biomedicines. 2022 Jul 28;10(8):1825. doi: 10.3390/biomedicines10081825. Biomedicines. 2022. PMID: 36009372 Free PMC article. Review.
References
-
- Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, Pulcini C, Kahlmeter G, Kluytmans J, Carmeli Y, Ouellette M, Outterson K, Patel J, Cavaleri M, Cox EM, Houchens CR, Grayson ML, Hansen P, Singh N, Theuretzbacher U, Magrini N, WHO Pathogens Priority List Working Group . 2018. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis 18:318–327. doi:10.1016/S1473-3099(17)30753-3. - DOI - PubMed
-
- Baltz R. 2007. Antimicrobials from actinomycetes: back to the future. Microbe 2:125–131.
-
- Mitcheltree MJ, Pisipati A, Syroegin EA, Silvestre KJ, Klepacki D, Mason JD, Terwilliger DW, Testolin G, Pote AR, Wu KJY, Ladley RP, Chatman K, Mankin AS, Polikanov YS, Myers AG. 2021. A synthetic antibiotic class overcoming bacterial multidrug resistance. Nature 599:507–512. doi:10.1038/s41586-021-04045-6. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
