

Representations of Temporal Community Structure in Hippocampus and Precuneus Predict Inductive Reasoning Decisions
- PMID: 35579986
- PMCID: PMC10262802
- DOI: 10.1162/jocn_a_01864
Representations of Temporal Community Structure in Hippocampus and Precuneus Predict Inductive Reasoning Decisions
Abstract
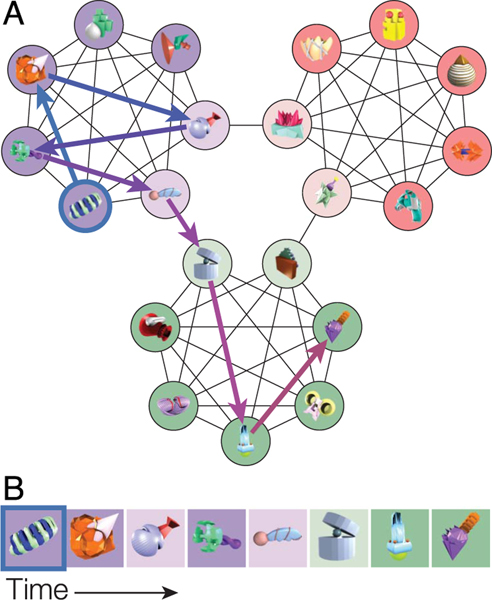
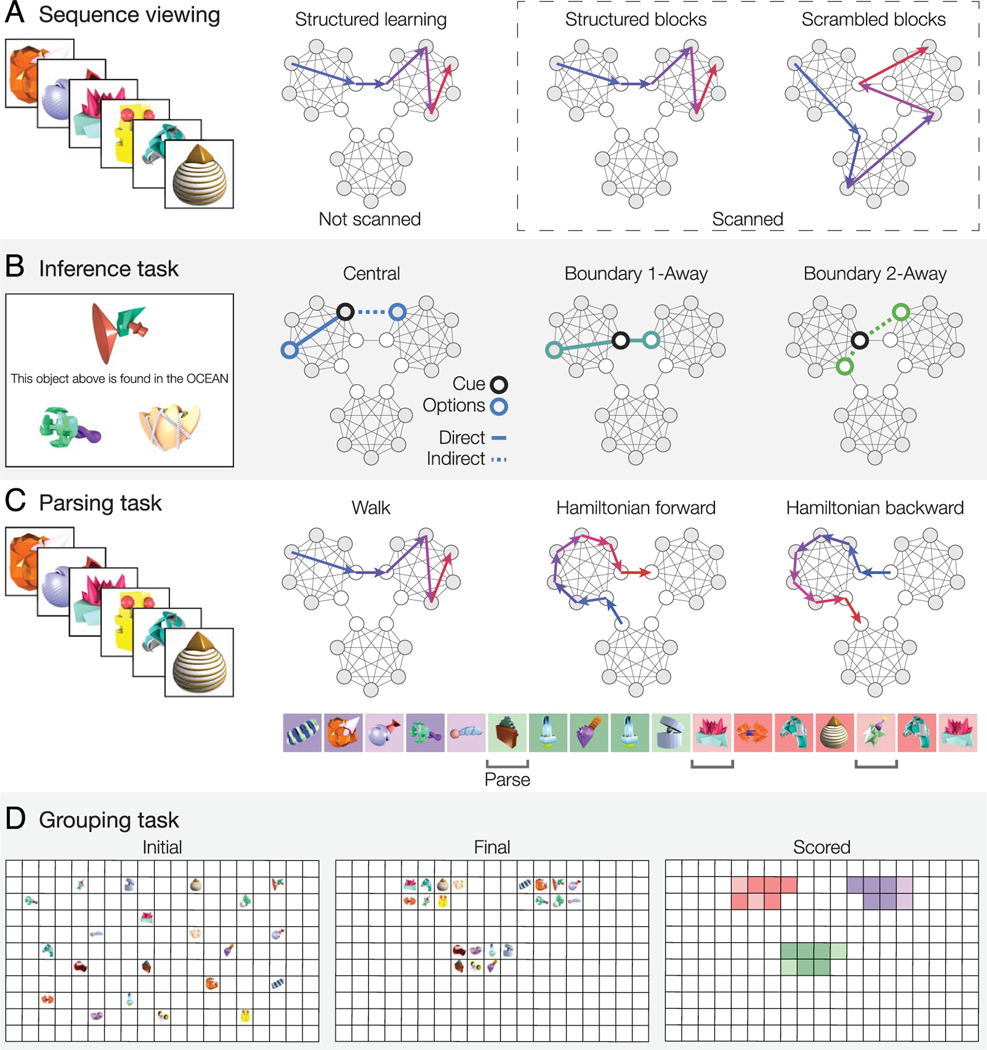
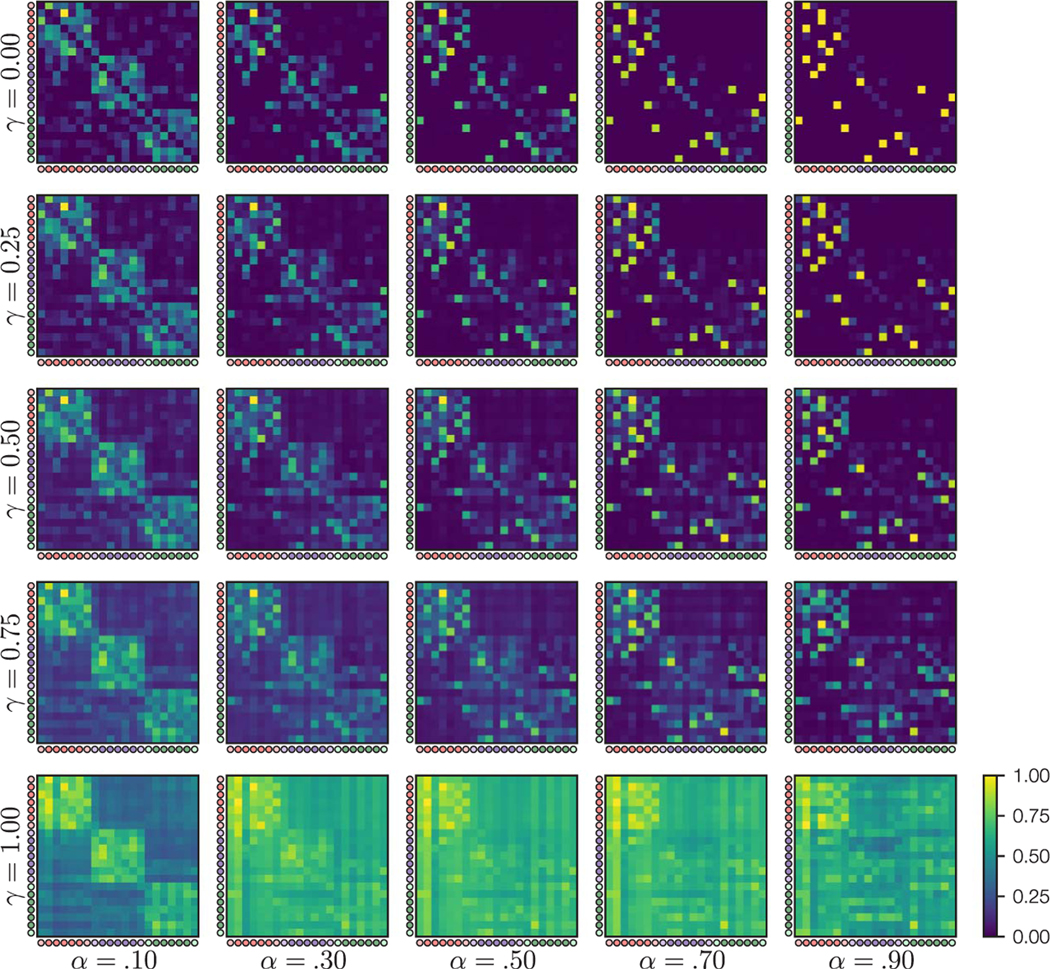
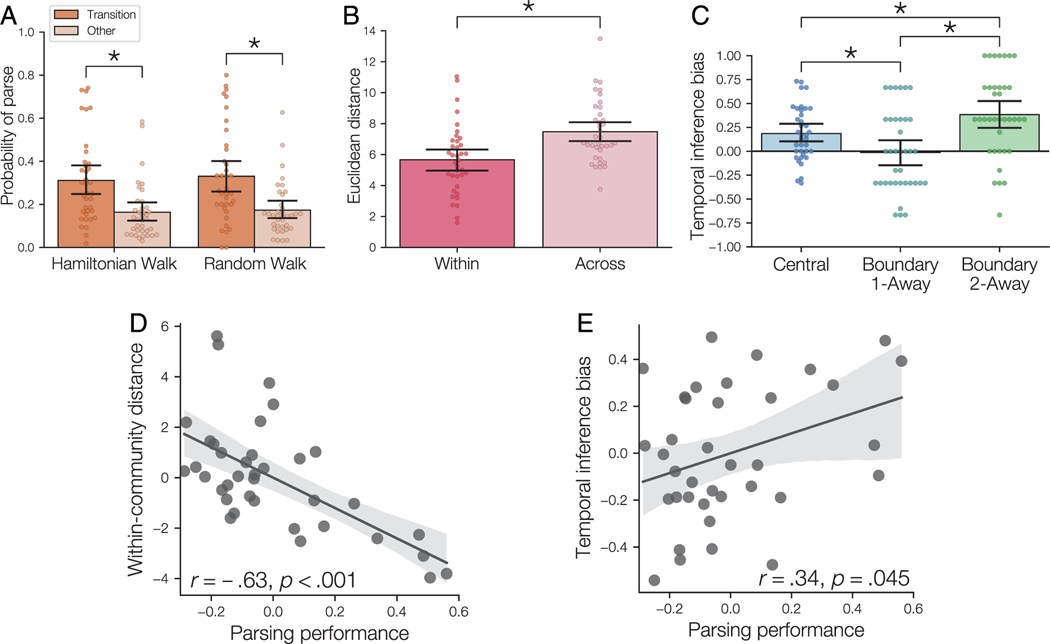
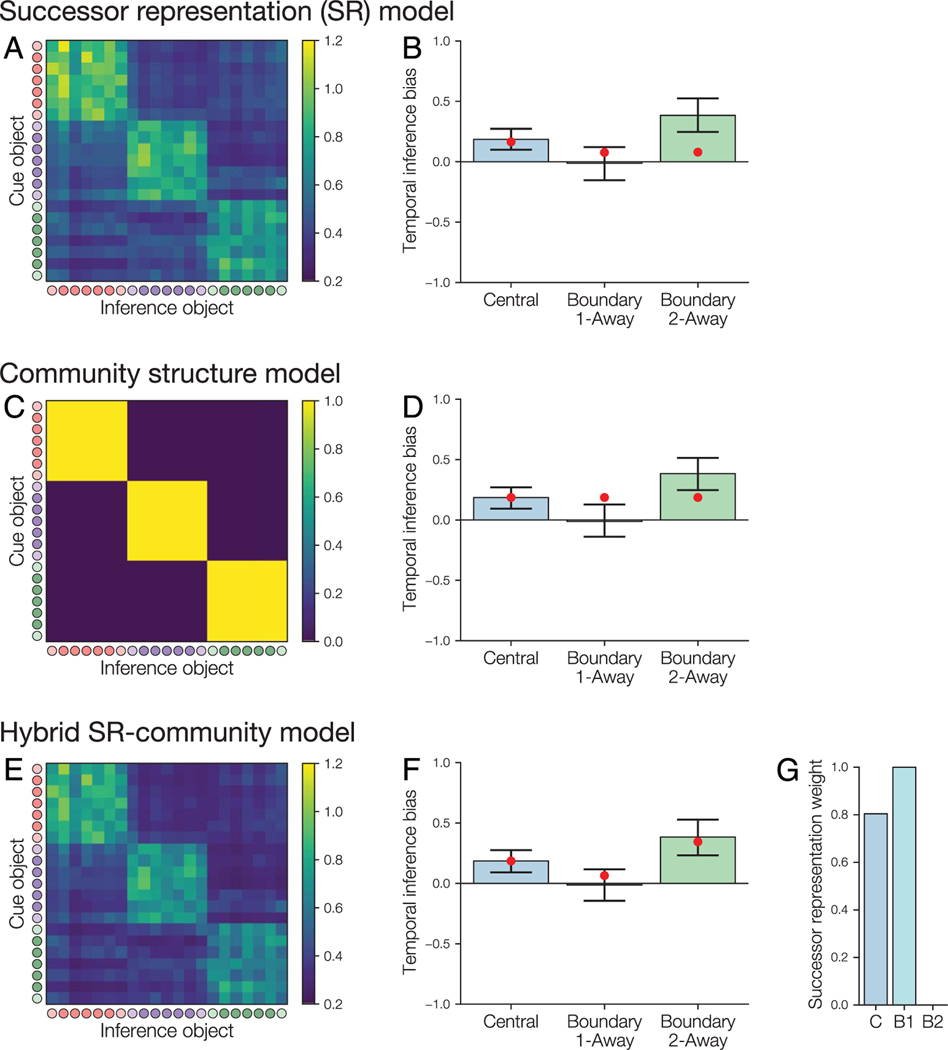
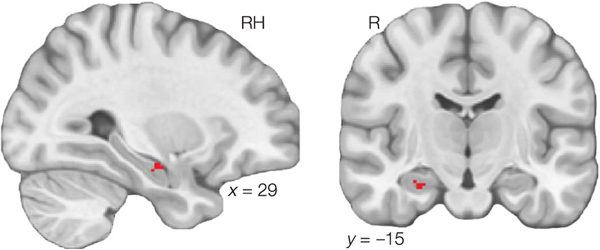
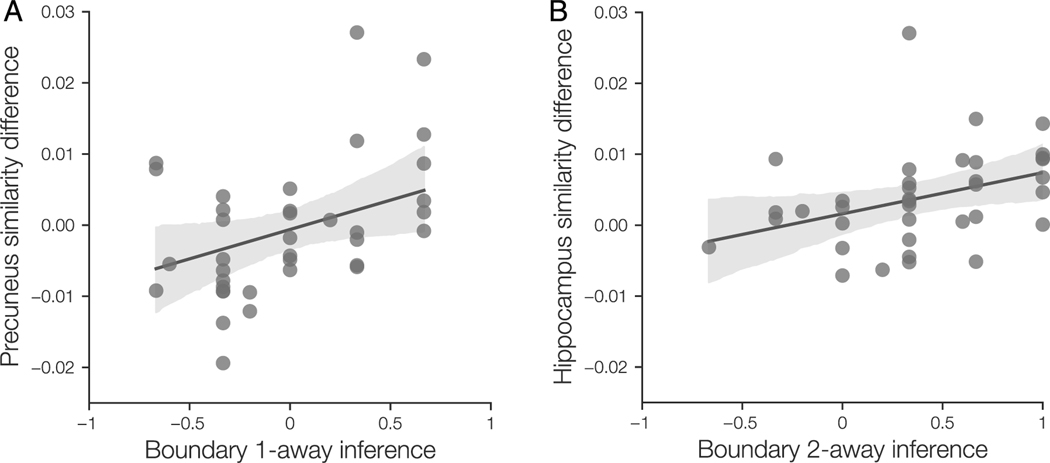
Our understanding of the world is shaped by inferences about underlying structure. For example, at the gym, you might notice that the same people tend to arrive around the same time and infer that they are friends that work out together. Consistent with this idea, after participants are presented with a temporal sequence of objects that follows an underlying community structure, they are biased to infer that objects from the same community share the same properties. Here, we used fMRI to measure neural representations of objects after temporal community structure learning and examine how these representations support inference about object relationships. We found that community structure learning affected inferred object similarity: When asked to spatially group items based on their experience, participants tended to group together objects from the same community. Neural representations in perirhinal cortex predicted individual differences in object grouping, suggesting that high-level object representations are affected by temporal community learning. Furthermore, participants were biased to infer that objects from the same community would share the same properties. Using computational modeling of temporal learning and inference decisions, we found that inductive reasoning is influenced by both detailed knowledge of temporal statistics and abstract knowledge of the temporal communities. The fidelity of temporal community representations in hippocampus and precuneus predicted the degree to which temporal community membership biased reasoning decisions. Our results suggest that temporal knowledge is represented at multiple levels of abstraction, and that perirhinal cortex, hippocampus, and precuneus may support inference based on this knowledge.
© 2022 Massachusetts Institute of Technology.
Figures

References
-
- Akaike H. (1998). Selected papers of Hirotugu Akaike. (Springer series in statistics, pp. 199–213). Springer. 10.1007/978-1-4612-1694-0_15 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources