

A loosened gating mechanism of RIG-I leads to autoimmune disorders
- PMID: 35580046
- PMCID: PMC9177982
- DOI: 10.1093/nar/gkac361
A loosened gating mechanism of RIG-I leads to autoimmune disorders
Abstract
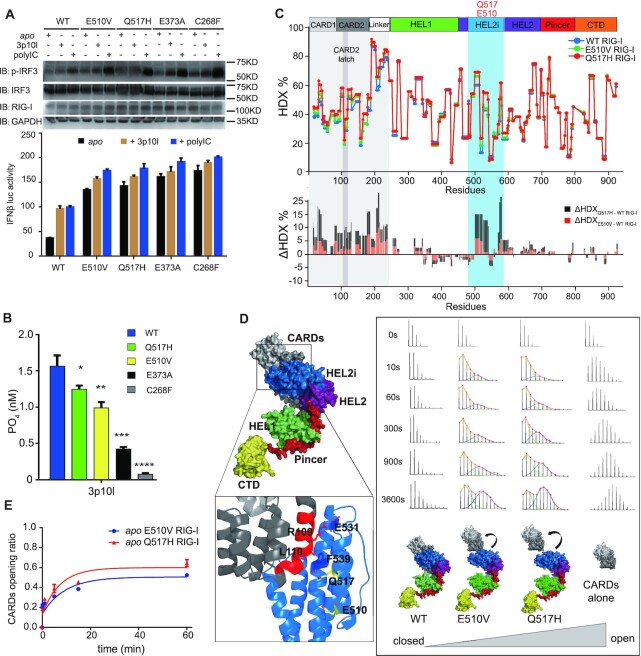
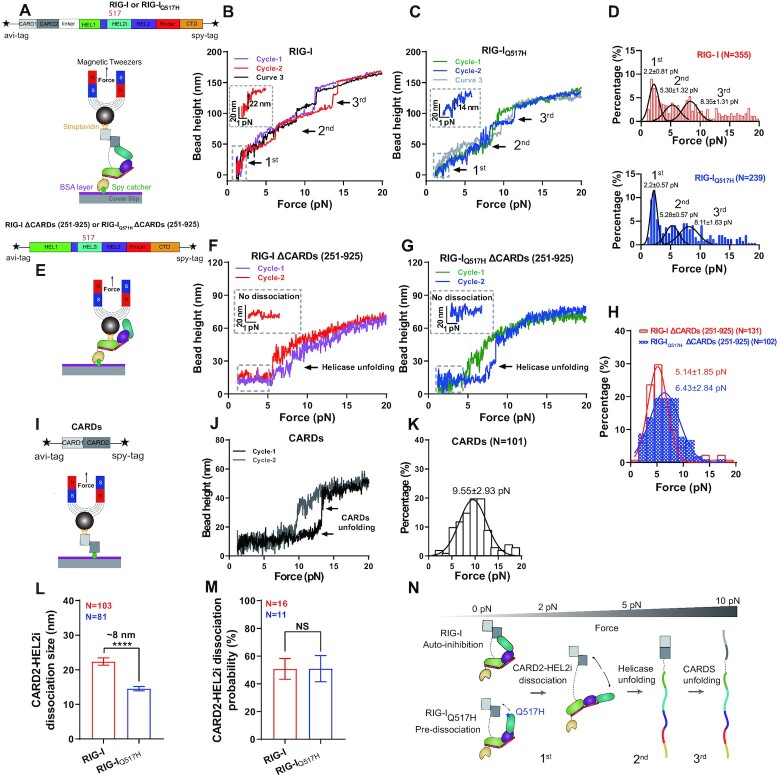
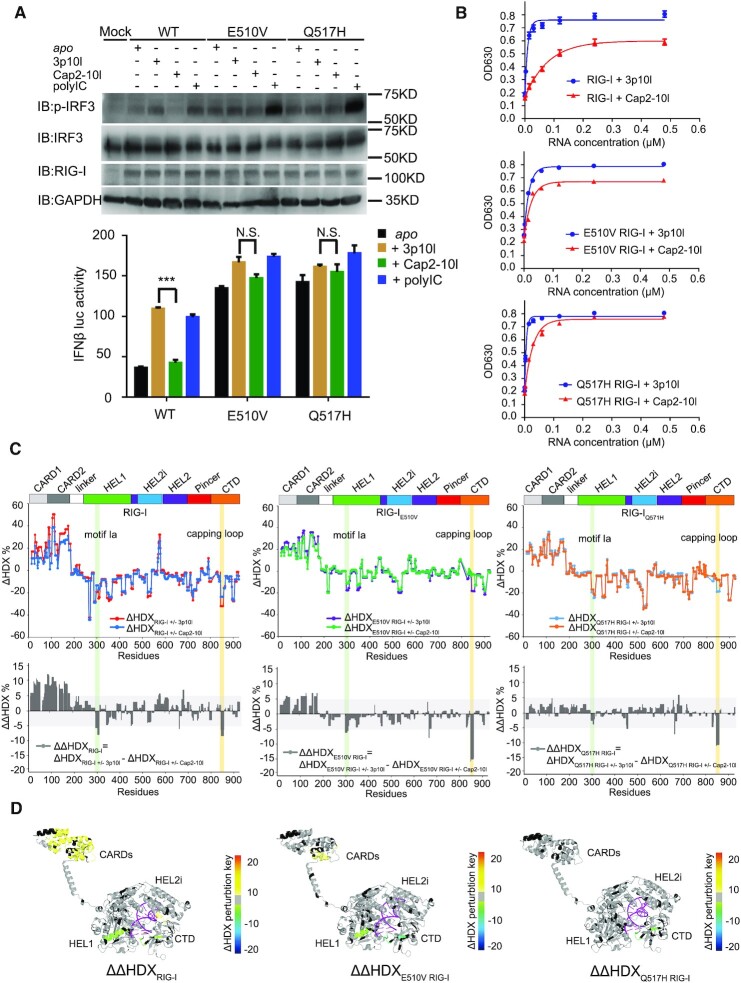
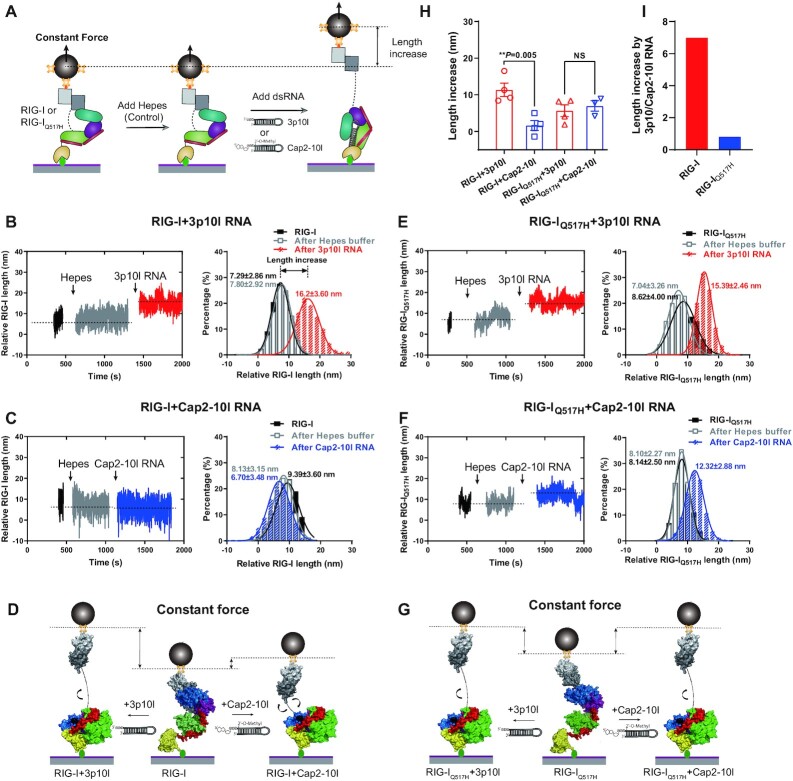
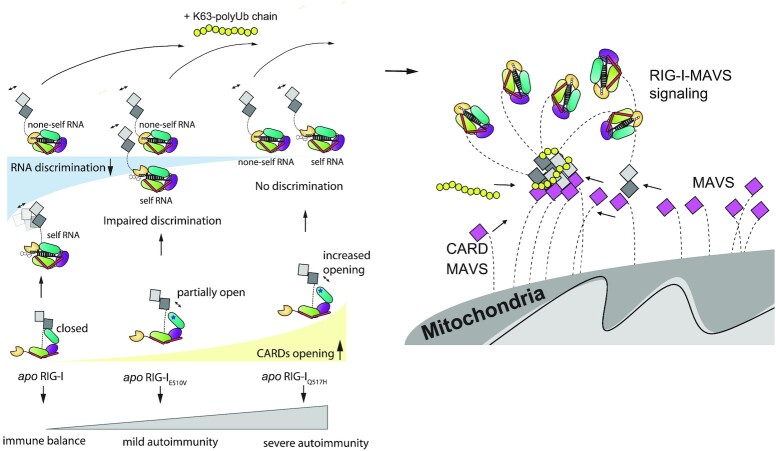
DDX58 encodes RIG-I, a cytosolic RNA sensor that ensures immune surveillance of nonself RNAs. Individuals with RIG-IE510V and RIG-IQ517H mutations have increased susceptibility to Singleton-Merten syndrome (SMS) defects, resulting in tissue-specific (mild) and classic (severe) phenotypes. The coupling between RNA recognition and conformational changes is central to RIG-I RNA proofreading, but the molecular determinants leading to dissociated disease phenotypes remain unknown. Herein, we employed hydrogen/deuterium exchange mass spectrometry (HDX-MS) and single molecule magnetic tweezers (MT) to precisely examine how subtle conformational changes in the helicase insertion domain (HEL2i) promote impaired ATPase and erroneous RNA proofreading activities. We showed that the mutations cause a loosened latch-gate engagement in apo RIG-I, which in turn gradually dampens its self RNA (Cap2 moiety:m7G cap and N1-2-2'-O-methylation RNA) proofreading ability, leading to increased immunopathy. These results reveal HEL2i as a unique checkpoint directing two specialized functions, i.e. stabilizing the CARD2-HEL2i interface and gating the helicase from incoming self RNAs; thus, these findings add new insights into the role of HEL2i in the control of antiviral innate immunity and autoimmunity diseases.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
The intrinsically disordered CARDs-Helicase linker in RIG-I is a molecular gate for RNA proofreading.EMBO J. 2022 May 16;41(10):e109782. doi: 10.15252/embj.2021109782. Epub 2022 Apr 19. EMBO J. 2022. PMID: 35437807 Free PMC article.
-
Unified mechanisms for self-RNA recognition by RIG-I Singleton-Merten syndrome variants.Elife. 2018 Jul 26;7:e38958. doi: 10.7554/eLife.38958. Elife. 2018. PMID: 30047865 Free PMC article.
-
HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity.Nat Commun. 2018 Dec 18;9(1):5366. doi: 10.1038/s41467-018-07780-z. Nat Commun. 2018. PMID: 30560918 Free PMC article.
-
What Really Rigs Up RIG-I?J Innate Immun. 2016;8(5):429-36. doi: 10.1159/000447947. Epub 2016 Jul 21. J Innate Immun. 2016. PMID: 27438016 Free PMC article. Review.
-
[Structural and functional views of the intracellular viral RNA sensor RIG-I].Uirusu. 2008 Dec;58(2):97-103. doi: 10.2222/jsv.58.97. Uirusu. 2008. PMID: 19374188 Review. Japanese.
Cited by
-
The m7G Reader NCBP2 Promotes Pancreatic Cancer Progression by Upregulating MAPK/ERK Signaling.Cancers (Basel). 2023 Nov 17;15(22):5454. doi: 10.3390/cancers15225454. Cancers (Basel). 2023. PMID: 38001714 Free PMC article.
-
Proofreading mechanisms of the innate immune receptor RIG-I: distinguishing self and viral RNA.Biochem Soc Trans. 2024 Jun 26;52(3):1131-1148. doi: 10.1042/BST20230724. Biochem Soc Trans. 2024. PMID: 38884803 Free PMC article. Review.
-
Sterile activation of RNA-sensing pathways in autoimmunity.J Mol Cell Biol. 2024 Dec 20;16(7):mjae029. doi: 10.1093/jmcb/mjae029. J Mol Cell Biol. 2024. PMID: 39143032 Free PMC article. Review.
-
Immunoengineering of a Photocaged 5´-triphosphate Oligoribonucleotide Ligand for Spatiotemporal Control of RIG-I Activation in Cancer.Angew Chem Int Ed Engl. 2025 May;64(21):e202423321. doi: 10.1002/anie.202423321. Epub 2025 Apr 21. Angew Chem Int Ed Engl. 2025. PMID: 40095771 Free PMC article.
-
DDX58 expression promotes inflammation and growth arrest in Sertoli cells by stabilizing p65 mRNA in patients with Sertoli cell-only syndrome.Front Immunol. 2023 Mar 22;14:1135753. doi: 10.3389/fimmu.2023.1135753. eCollection 2023. Front Immunol. 2023. PMID: 37033952 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
