

Structural insights into the HBV receptor and bile acid transporter NTCP
- PMID: 35580630
- PMCID: PMC9242859
- DOI: 10.1038/s41586-022-04857-0
Structural insights into the HBV receptor and bile acid transporter NTCP
Abstract
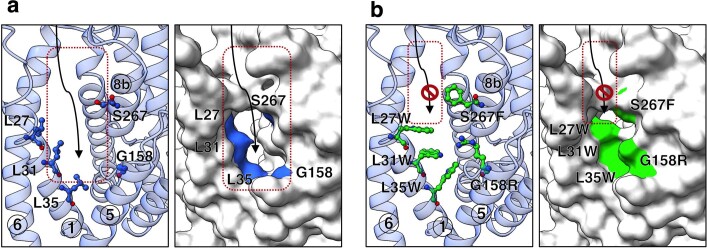
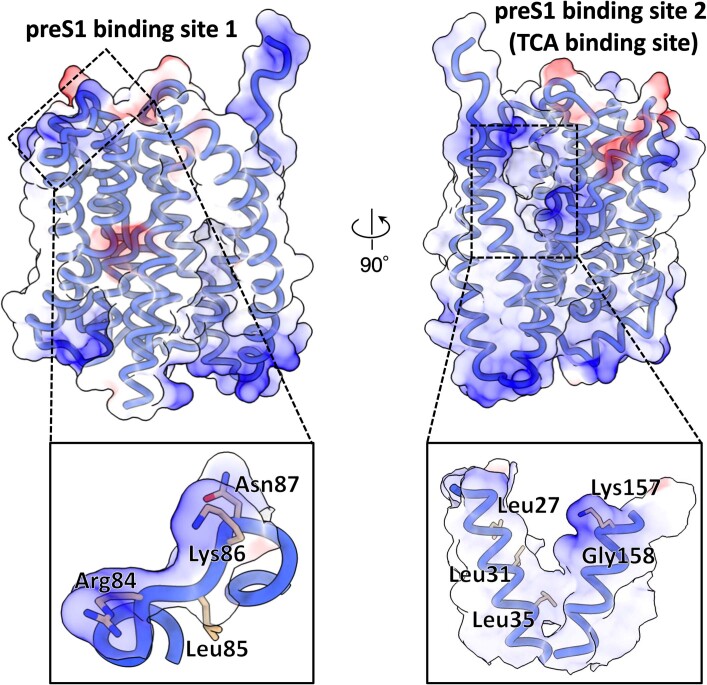
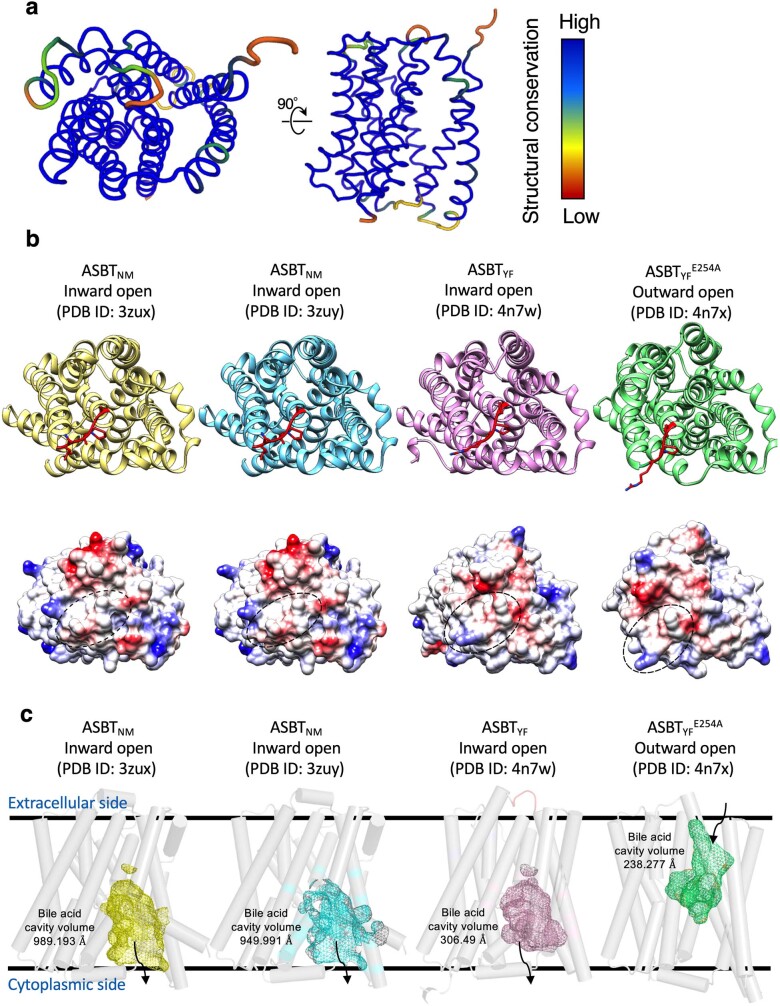
Around 250 million people are infected with hepatitis B virus (HBV) worldwide1, and 15 million may also carry the satellite virus hepatitis D virus (HDV), which confers even greater risk of severe liver disease2. The HBV receptor has been identified as sodium taurocholate co-transporting polypeptide (NTCP), which interacts directly with the first 48 amino acid residues of the N-myristoylated N-terminal preS1 domain of the viral large protein3. Despite the pressing need for therapeutic agents to counter HBV, the structure of NTCP remains unsolved. This 349-residue protein is closely related to human apical sodium-dependent bile acid transporter (ASBT), another member of the solute carrier family SLC10. Crystal structures have been reported of similar bile acid transporters from bacteria4,5, and these models are believed to resemble closely both NTCP and ASBT. Here we have used cryo-electron microscopy to solve the structure of NTCP bound to an antibody, clearly showing that the transporter has no equivalent of the first transmembrane helix found in other SLC10 proteins, and that the N terminus is exposed on the extracellular face. Comparison of our structure with those of related proteins indicates a common mechanism of bile acid transport, but the NTCP structure displays an additional pocket formed by residues that are known to interact with preS1, presenting new opportunities for structure-based drug design.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
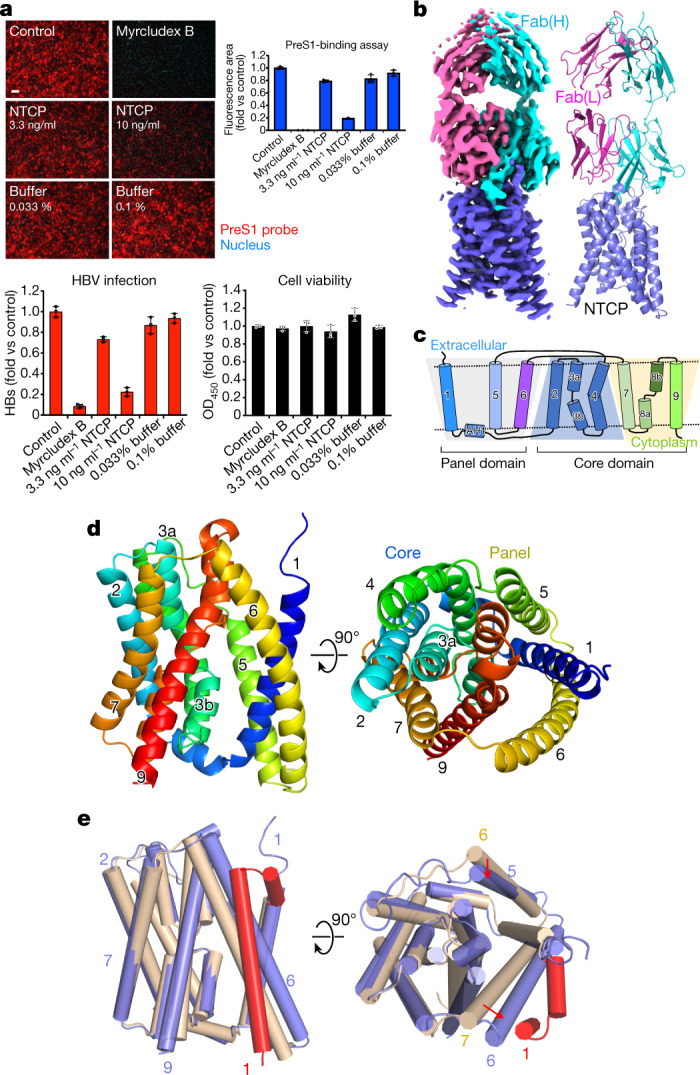
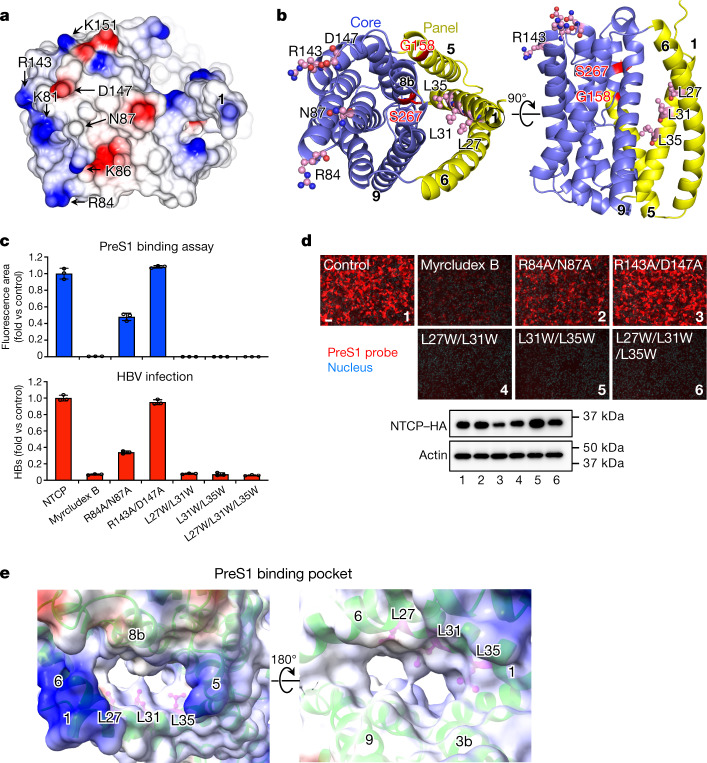
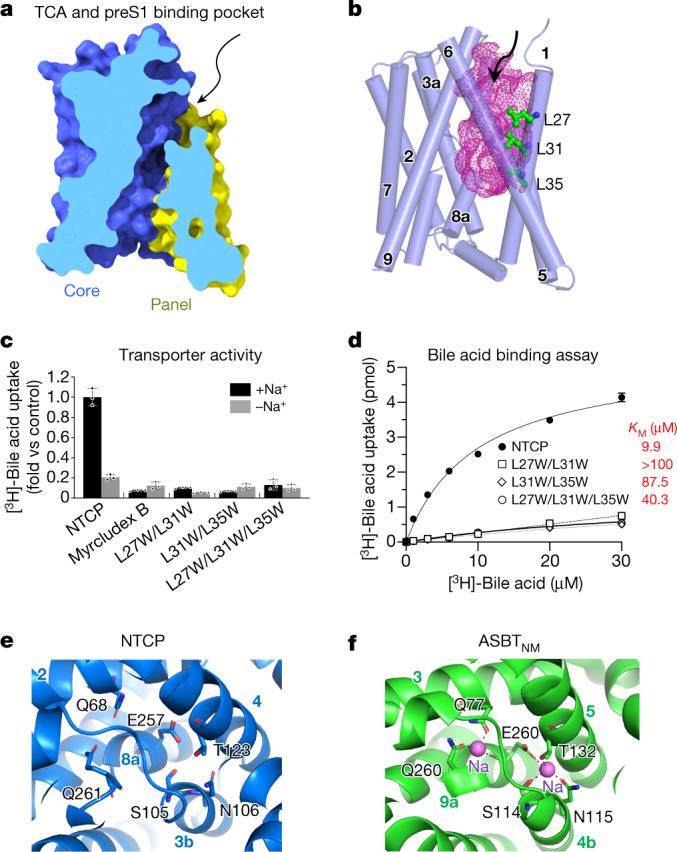
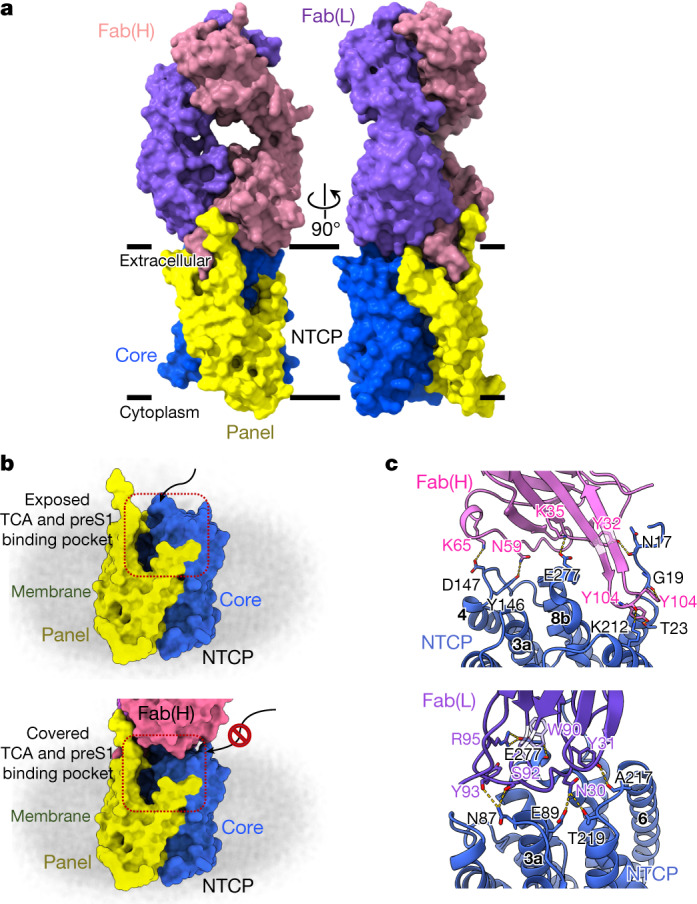

Similar articles
-
Structural basis of sodium-dependent bile salt uptake into the liver.Nature. 2022 Jun;606(7916):1015-1020. doi: 10.1038/s41586-022-04723-z. Epub 2022 May 11. Nature. 2022. PMID: 35545671 Free PMC article.
-
Viral entry of hepatitis B and D viruses and bile salts transportation share common molecular determinants on sodium taurocholate cotransporting polypeptide.J Virol. 2014 Mar;88(6):3273-84. doi: 10.1128/JVI.03478-13. Epub 2014 Jan 3. J Virol. 2014. PMID: 24390325 Free PMC article.
-
Structure of the bile acid transporter and HBV receptor NTCP.Nature. 2022 Jun;606(7916):1021-1026. doi: 10.1038/s41586-022-04845-4. Epub 2022 May 17. Nature. 2022. PMID: 35580629
-
Molecular regulation of the hepatic bile acid uptake transporter and HBV entry receptor NTCP.Biochim Biophys Acta Mol Cell Biol Lipids. 2021 Aug;1866(8):158960. doi: 10.1016/j.bbalip.2021.158960. Epub 2021 Apr 29. Biochim Biophys Acta Mol Cell Biol Lipids. 2021. PMID: 33932583 Review.
-
Innovative HBV Animal Models Based on the Entry Receptor NTCP.Viruses. 2020 Jul 30;12(8):828. doi: 10.3390/v12080828. Viruses. 2020. PMID: 32751581 Free PMC article. Review.
Cited by
-
Tyrosine 146 of the Human Na+/Taurocholate Cotransporting Polypeptide (NTCP) Is Essential for Its Hepatitis B Virus (HBV) Receptor Function and HBV Entry into Hepatocytes.Viruses. 2022 Jun 9;14(6):1259. doi: 10.3390/v14061259. Viruses. 2022. PMID: 35746730 Free PMC article.
-
HBV Vaccines: Advances and Development.Vaccines (Basel). 2023 Dec 18;11(12):1862. doi: 10.3390/vaccines11121862. Vaccines (Basel). 2023. PMID: 38140265 Free PMC article. Review.
-
Recent Advances in mRNA-Based Vaccines Against Several Hepatitis Viruses.Biol Proced Online. 2025 Jun 3;27(1):20. doi: 10.1186/s12575-025-00269-2. Biol Proced Online. 2025. PMID: 40461976 Free PMC article. Review.
-
Structural basis of hepatitis B virus receptor binding.Nat Struct Mol Biol. 2024 Mar;31(3):447-454. doi: 10.1038/s41594-023-01191-5. Epub 2024 Jan 17. Nat Struct Mol Biol. 2024. PMID: 38233573
-
Role of the Sodium-Dependent Organic Anion Transporter (SOAT/SLC10A6) in Physiology and Pathophysiology.Int J Mol Sci. 2023 Jun 8;24(12):9926. doi: 10.3390/ijms24129926. Int J Mol Sci. 2023. PMID: 37373074 Free PMC article. Review.
References
-
- Global Hepatitis Report (World Health Organization, 2017).
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
