

The K-Ras(G12D)-inhibitory peptide KS-58 suppresses growth of murine CT26 colorectal cancer cell-derived tumors
- PMID: 35581303
- PMCID: PMC9114382
- DOI: 10.1038/s41598-022-12401-3
The K-Ras(G12D)-inhibitory peptide KS-58 suppresses growth of murine CT26 colorectal cancer cell-derived tumors
Abstract
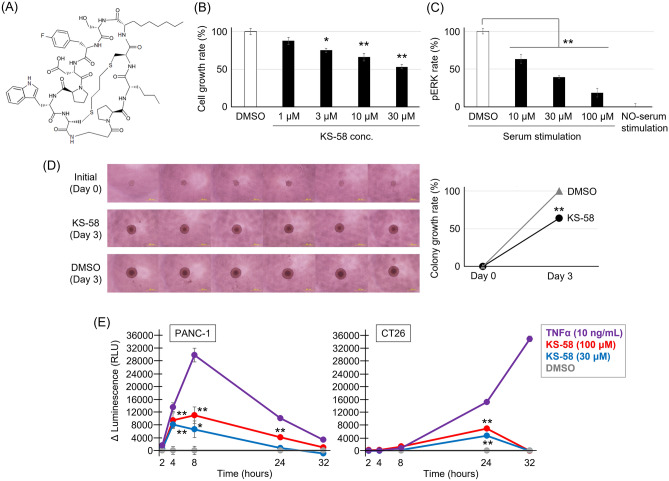
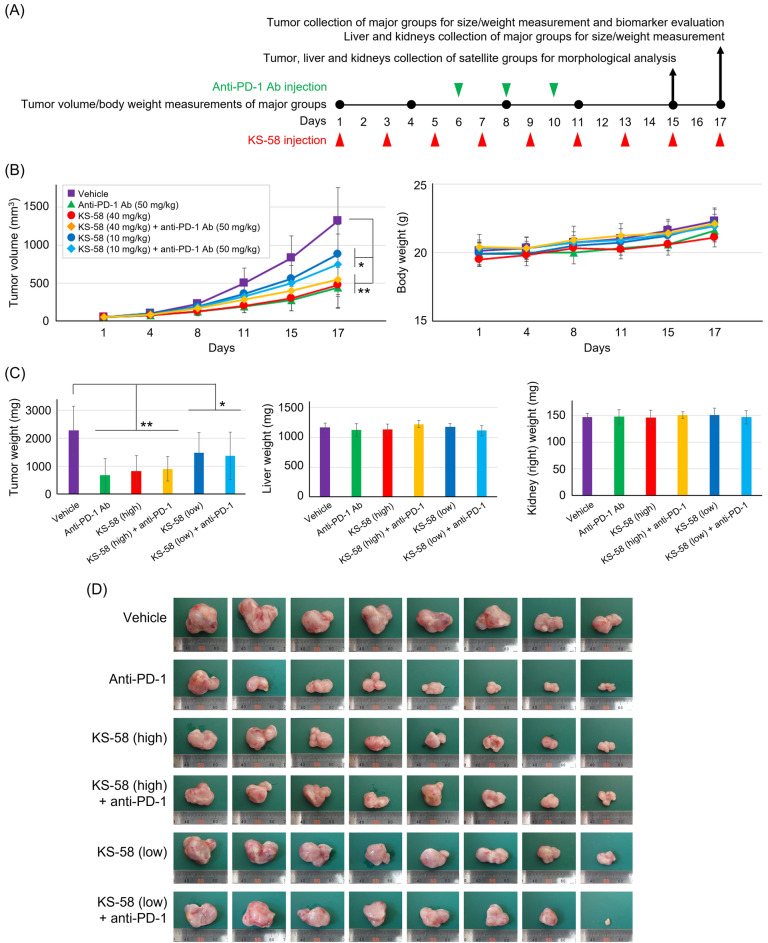
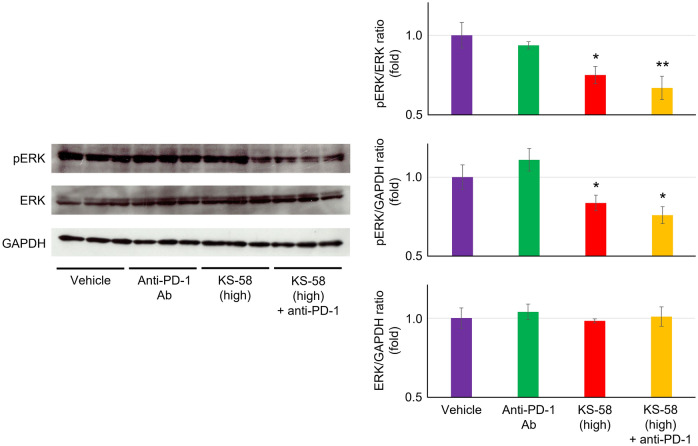
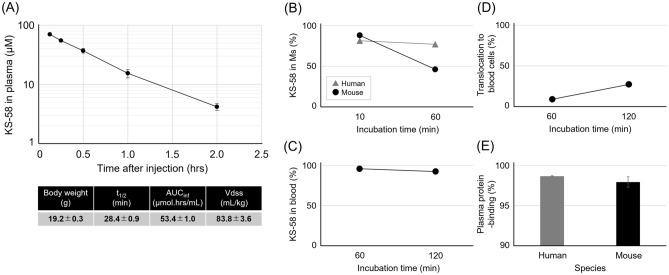
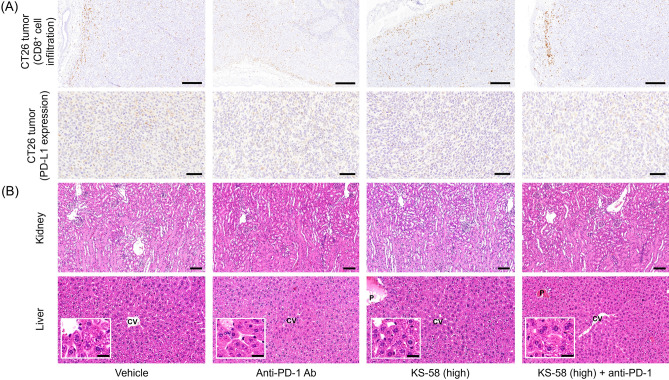
Mutations in the cell proliferation regulator K-Ras are found with a variety of cancer types, so drugs targeting these mutant proteins could hold great clinical potential. Very recently, a drug targeting the K-Ras(G12C) mutant observed in lung cancer gained regulatory approval and several clinical trials are currently underway to examine the efficacy of this agent when combined with other drugs such as a monoclonal antibody inhibitor of programmed cell death 1 receptor (anti-PD-1). Alternatively, there are currently no approved drugs targeting K-Ras(G12D), the most common cancer-associated K-Ras mutant. In 2020, we described the development of the K-Ras(G12D) inhibitory bicyclic peptide KS-58 and presented evidence for anticancer activity against mouse xenografts derived from the human pancreatic cancer cell line PANC-1 stably expressing K-Ras(G12D). Here, we show that KS-58 also possess anticancer activity against mouse tumors derived from the colorectal cancer cell line CT26 stably expressing K-Ras(G12D). Further, KS-58 treatment reduced phosphorylation of ERK, a major downstream signaling factor in the Ras pathway, confirming that KS-58 inhibits K-Ras(G12D) function. Unexpectedly; however, KS-58 did not show additive or synergistic anticancer activity with mouse anti-PD-1. Morphological analysis and immunostaining demonstrated no obvious differences in CD8+ cells infiltration or PD-L1 expression levels in CT26-derived tumors exposed to monotherapy or combination treatment. Nonetheless, KS-58 demonstrated reasonable stability in blood (t1/2 ≈ 30 min) and no obvious systemic adverse effects, suggesting clinical potential as a lead molecule against colorectal cancer.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Nanoformulation of the K-Ras(G12D)-inhibitory peptide KS-58 suppresses colorectal and pancreatic cancer-derived tumors.Sci Rep. 2023 Jan 10;13(1):518. doi: 10.1038/s41598-023-27825-8. Sci Rep. 2023. PMID: 36627374 Free PMC article.
-
Generation of KS-58 as the first K-Ras(G12D)-inhibitory peptide presenting anti-cancer activity in vivo.Sci Rep. 2020 Dec 10;10(1):21671. doi: 10.1038/s41598-020-78712-5. Sci Rep. 2020. PMID: 33303890 Free PMC article.
-
Feedback activation of EGFR/wild-type RAS signaling axis limits KRASG12D inhibitor efficacy in KRASG12D-mutated colorectal cancer.Oncogene. 2023 May;42(20):1620-1633. doi: 10.1038/s41388-023-02676-9. Epub 2023 Apr 5. Oncogene. 2023. PMID: 37020035 Free PMC article.
-
Targeting Krasg12c -mutant cancer with a mutation-specific inhibitor.J Intern Med. 2020 Aug;288(2):183-191. doi: 10.1111/joim.13057. Epub 2020 Apr 7. J Intern Med. 2020. PMID: 32176377 Review.
-
Peptides That Block RAS-p21 Protein-Induced Cell Transformation.Biomedicines. 2023 Feb 6;11(2):471. doi: 10.3390/biomedicines11020471. Biomedicines. 2023. PMID: 36831007 Free PMC article. Review.
Cited by
-
A FEN 1-driven DNA walker-like reaction coupling with magnetic bead-based separation for specific SNP detection.Front Bioeng Biotechnol. 2023 Nov 3;11:1279473. doi: 10.3389/fbioe.2023.1279473. eCollection 2023. Front Bioeng Biotechnol. 2023. PMID: 38026850 Free PMC article.
-
The Nanotechnology-Based Approaches against Kirsten Rat Sarcoma-Mutated Cancers.Pharmaceutics. 2023 Jun 8;15(6):1686. doi: 10.3390/pharmaceutics15061686. Pharmaceutics. 2023. PMID: 37376135 Free PMC article. Review.
-
Nanoformulation of the K-Ras(G12D)-inhibitory peptide KS-58 suppresses colorectal and pancreatic cancer-derived tumors.Sci Rep. 2023 Jan 10;13(1):518. doi: 10.1038/s41598-023-27825-8. Sci Rep. 2023. PMID: 36627374 Free PMC article.
-
Experimental Murine Models for Colorectal Cancer Research.Cancers (Basel). 2023 Apr 30;15(9):2570. doi: 10.3390/cancers15092570. Cancers (Basel). 2023. PMID: 37174036 Free PMC article. Review.
-
Driver Mutations in Pancreatic Cancer and Opportunities for Targeted Therapy.Cancers (Basel). 2024 May 9;16(10):1808. doi: 10.3390/cancers16101808. Cancers (Basel). 2024. PMID: 38791887 Free PMC article. Review.
References
-
- Canon J, Rex K, Saiki YA, Mohr C, Cooke K, Bagal D, Gaida K, Holt T, Knutson GC, Koppada N, Lanman AB, Werner J, Rapaport SA, Miguel ST, Ortiz R, Osgood T, Sun JR, Zhu X, McCarter DJ, Volak PL, Houk EB, Fakih GM, O’Neil HB, Price JT, Falchook SG, Desai J, Kuo J, Govindan R, Hong SD, Ouyang W, Henary H, Arvedson T, Cee JV, Lipford RJ. The clinical KRAS(G12C) inhibitor amg 510 drives anti-tumor immunity. Nature. 2019;575:217–223. doi: 10.1038/s41586-019-1694-1. - DOI - PubMed
-
- Briere MD, Li S, Calinisan A, Sudhakar N, Aranda R, Hargis L, Hargis L, Peng HD, Geng J, Engstrom DL, Hallin J, Gatto S, Fernandez-Banet J, Pavlicek A, Wong KK, Christensen GJ, Olson P. The KRASG12C Inhibitor MRTX849 reconditions the tumor immune microenvironment and sensitizes tumors to checkpoint inhibitor therapy. Mol. Cancer Ther. 2021;20:975–985. doi: 10.1158/1535-7163.MCT-20-0462. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
