

Identification of LINE retrotransposons and long non-coding RNAs expressed in the octopus brain
- PMID: 35581640
- PMCID: PMC9115989
- DOI: 10.1186/s12915-022-01303-5
Identification of LINE retrotransposons and long non-coding RNAs expressed in the octopus brain
Abstract
Background: Transposable elements (TEs) widely contribute to the evolution of genomes allowing genomic innovations, generating germinal and somatic heterogeneity, and giving birth to long non-coding RNAs (lncRNAs). These features have been associated to the evolution, functioning, and complexity of the nervous system at such a level that somatic retrotransposition of long interspersed element (LINE) L1 has been proposed to be associated to human cognition. Among invertebrates, octopuses are fascinating animals whose nervous system reaches a high level of complexity achieving sophisticated cognitive abilities. The sequencing of the genome of the Octopus bimaculoides revealed a striking expansion of TEs which were proposed to have contributed to the evolution of its complex nervous system. We recently found a similar expansion also in the genome of Octopus vulgaris. However, a specific search for the existence and the transcription of full-length transpositionally competent TEs has not been performed in this genus.
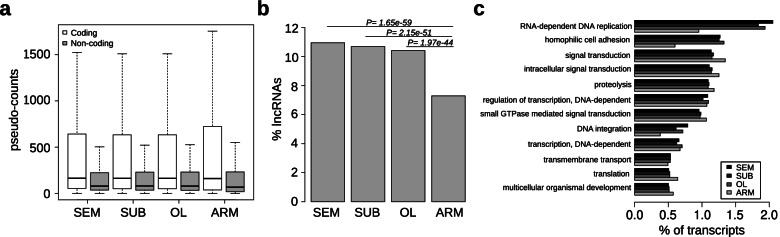
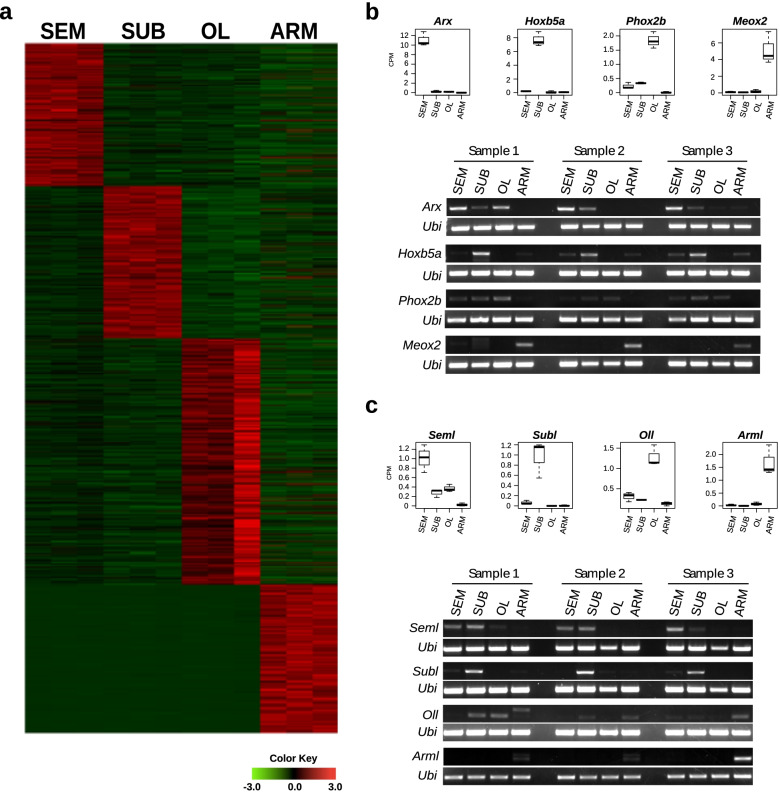
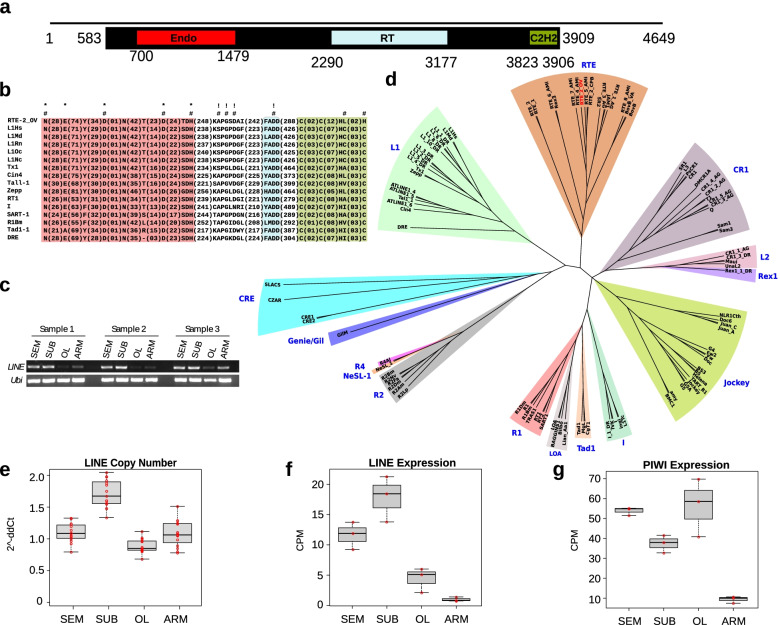
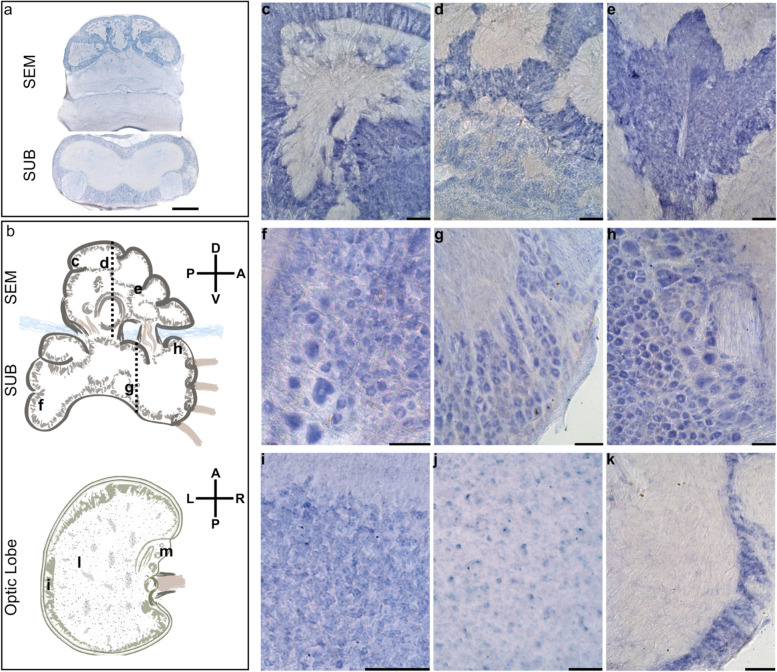
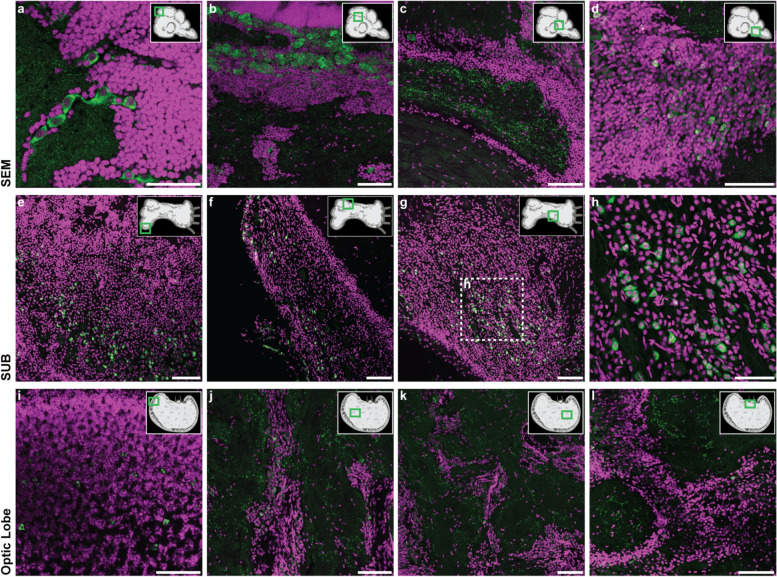
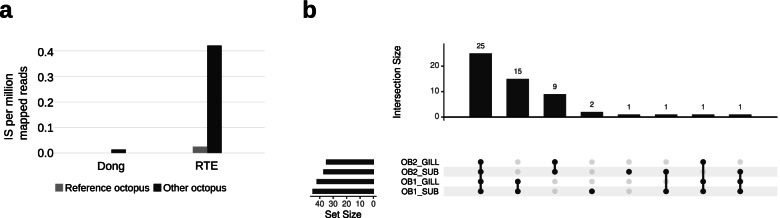
Results: Here, we report the identification of LINE elements competent for retrotransposition in Octopus vulgaris and Octopus bimaculoides and show evidence suggesting that they might be transcribed and determine germline and somatic polymorphisms especially in the brain. Transcription and translation measured for one of these elements resulted in specific signals in neurons belonging to areas associated with behavioral plasticity. We also report the transcription of thousands of lncRNAs and the pervasive inclusion of TE fragments in the transcriptomes of both Octopus species, further testifying the crucial activity of TEs in the evolution of the octopus genomes.
Conclusions: The neural transcriptome of the octopus shows the transcription of thousands of putative lncRNAs and of a full-length LINE element belonging to the RTE class. We speculate that a convergent evolutionary process involving retrotransposons activity in the brain has been important for the evolution of sophisticated cognitive abilities in this genus.
Keywords: Mollusks; Nervous system; Transcriptome; Transposable elements.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
A chromosome-level reference genome for the common octopus, Octopus vulgaris (Cuvier, 1797).G3 (Bethesda). 2023 Dec 6;13(12):jkad220. doi: 10.1093/g3journal/jkad220. G3 (Bethesda). 2023. PMID: 37850903 Free PMC article.
-
From the genome's perspective: Bearing somatic retrotransposition to leverage the regulatory potential of L1 RNAs.Bioessays. 2025 Feb;47(2):e2400125. doi: 10.1002/bies.202400125. Epub 2024 Nov 9. Bioessays. 2025. PMID: 39520370 Free PMC article. Review.
-
The octopus genome and the evolution of cephalopod neural and morphological novelties.Nature. 2015 Aug 13;524(7564):220-4. doi: 10.1038/nature14668. Nature. 2015. PMID: 26268193 Free PMC article.
-
Comparative analysis of transposable elements provides insights into genome evolution in the genus Camelus.BMC Genomics. 2021 Nov 20;22(1):842. doi: 10.1186/s12864-021-08117-9. BMC Genomics. 2021. PMID: 34800971 Free PMC article.
-
Mammalian transposable elements and their impacts on genome evolution.Chromosome Res. 2018 Mar;26(1-2):25-43. doi: 10.1007/s10577-017-9570-z. Epub 2018 Feb 1. Chromosome Res. 2018. PMID: 29392473 Free PMC article. Review.
Cited by
-
Epigenetic machinery is functionally conserved in cephalopods.BMC Biol. 2022 Sep 14;20(1):202. doi: 10.1186/s12915-022-01404-1. BMC Biol. 2022. PMID: 36104784 Free PMC article.
-
A chromosome-level reference genome for the common octopus, Octopus vulgaris (Cuvier, 1797).G3 (Bethesda). 2023 Dec 6;13(12):jkad220. doi: 10.1093/g3journal/jkad220. G3 (Bethesda). 2023. PMID: 37850903 Free PMC article.
-
From the genome's perspective: Bearing somatic retrotransposition to leverage the regulatory potential of L1 RNAs.Bioessays. 2025 Feb;47(2):e2400125. doi: 10.1002/bies.202400125. Epub 2024 Nov 9. Bioessays. 2025. PMID: 39520370 Free PMC article. Review.
-
Cell type diversity in a developing octopus brain.Nat Commun. 2022 Nov 30;13(1):7392. doi: 10.1038/s41467-022-35198-1. Nat Commun. 2022. PMID: 36450803 Free PMC article.
-
Transcriptome-wide selection and validation of a solid set of reference genes for gene expression studies in the cephalopod mollusk Octopus vulgaris.Front Mol Neurosci. 2023 May 17;16:1091305. doi: 10.3389/fnmol.2023.1091305. eCollection 2023. Front Mol Neurosci. 2023. PMID: 37266373 Free PMC article.
