

Characterization of uridine-cytidine kinase like-1 nucleoside kinase activity and its role in tumor growth
- PMID: 35583288
- PMCID: PMC9246348
- DOI: 10.1042/BCJ20210770
Characterization of uridine-cytidine kinase like-1 nucleoside kinase activity and its role in tumor growth
Abstract
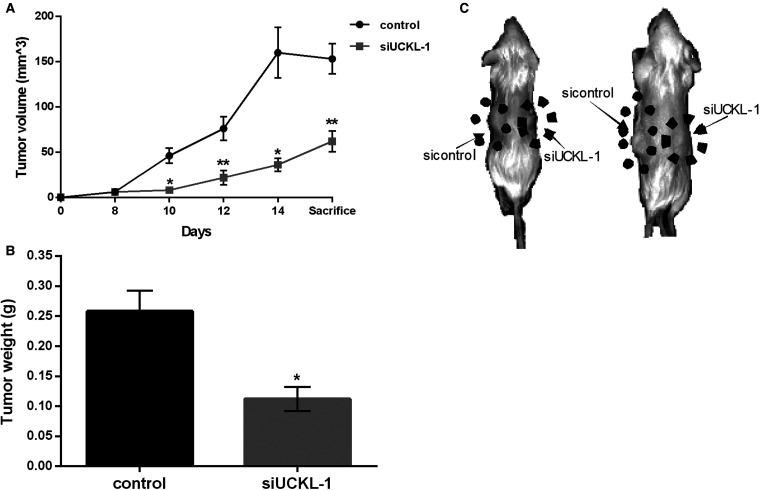
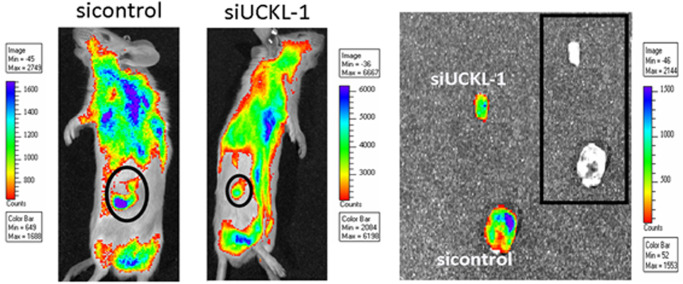
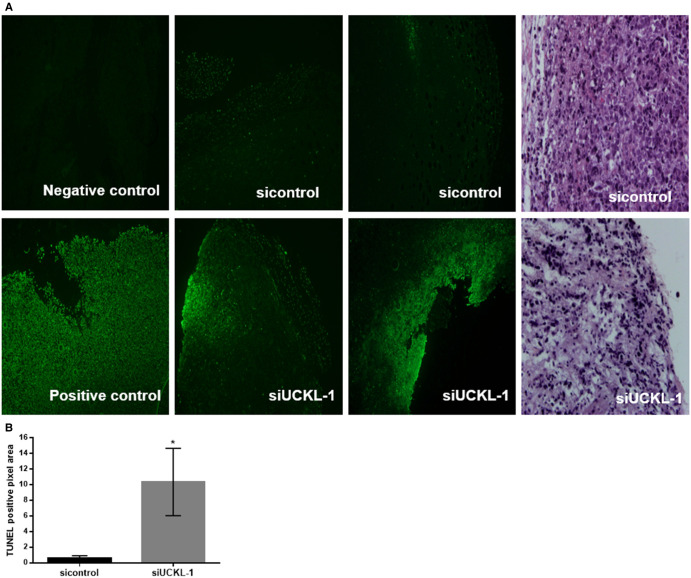
Uridine-cytidine kinase like-1 (UCKL-1) is a largely uncharacterized protein with high sequence similarity to other uridine-cytidine kinases (UCKs). UCKs play an important role in the pyrimidine salvage pathway, catalyzing the phosphorylation of uridine and cytidine to UMP and CMP, respectively. Only two human UCKs have been identified, UCK1 and UCK2. Previous studies have shown both enzymes phosphorylate uridine and cytidine using ATP as the phosphate donor. No studies have evaluated the kinase potential of UCKL-1. We cloned and purified UCKL-1 and found that it successfully phosphorylated uridine and cytidine using ATP as the phosphate donor. The catalytic efficiency (calculated as kcat/KM) was 1.2 × 104 s-1, M-1 for uridine and 0.7 × 104 s-1, M-1 for cytidine. Our lab has previously shown that UCKL-1 is up-regulated in tumor cells, providing protection against natural killer (NK) cell killing activity. We utilized small interfering RNA (siRNA) to down-regulate UCKL-1 in vitro and in vivo to determine the effect of UCKL-1 on tumor growth and metastasis. The down-regulation of UCKL-1 in YAC-1 lymphoma cells in vitro resulted in decreased cell counts and increased apoptotic activity. Down-regulation of UCKL-1 in K562 leukemia cells in vivo led to decreased primary tumor growth and less tumor cell dissemination and metastasis. These results identify UCKL-1 as a bona fide pyrimidine kinase with the therapeutic potential to be a target for tumor growth inhibition and for diminishing or preventing metastasis.
Keywords: cancer; metastasis; natural killer; nucleoside kinase; uridine kinase.
© 2022 The Author(s).
Conflict of interest statement
The authors declare that there are no competing interests associated with the manuscript.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous